

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Prostate cancer is the most common non-skin cancer and the second leading cause of cancer mortality in the male population (1). Recently, new targeted gene therapy using RNA interference (RNAi) is being investigated for the treatment of advanced prostate cancer. The RNAi relies on post-transcriptional gene silencing using double-strand RNA processed into 21–25 nucleotide length, known as small interfering RNA (siRNA) (2). When siRNA is incorporated into RNA-induced silencing complexes in the cytoplasm, it promotes targeted gene silencing by sequence-specific degradation of messenger RNA (23). The siRNA is a promising therapeutic approach for cancer treatment, since it can prevent the production of specific proteins essential for the proliferation of tumor cells.
Many approaches have been studied to enhance the delivery of treatment agents such as siRNA into tumor cells (456). Our study group developed microbubble-liposome complex (MLC) for the guided delivery of agents to the prostate cancer cells (7). Microbubbles, gas bubbles 1–8 µm in diameter (8910), can act as cavitation nuclei that carry drugs or genes to target cells, enabling site-targeted treatment (8910). Further application of ultrasound can facilitate microbubble-mediated delivery, by the collapse of microbubbles, perforations in cell membranes, and increased permeability of regional capillaries, which allows large molecules to stream into the cells (9). This phenomenon, called sonoporation, can expedite the intracellular ingress of drugs or genes in ultrasound microbubble-mediated delivery (911). Additionally, we linked liposomes with microbubbles to carry not only siRNA but also chemotherapeutic drug–doxorubicin. In a novel approach, we conjugated anti-human epidermal growth factor receptor type 2 (Her2) antibodies with MLC to target prostate cancer cells that are known to express Her2 (12).
The purpose of our study was to evaluate the effectiveness of ultrasound and MLC-mediated delivery of siRNA and doxorubicin into prostate cancer cells and the therapeutic capability of this approach both in vitro and in vivo.
MATERIALS AND METHODS
Cell Lines and Culture
Human prostate cancer cell lines PC-3 and LNCaP were obtained from American Type Culture Collection (Manassas, VA, USA). Cells were maintained in RPMI-1640 medium supplemented with 10% fetal bovine serum at 37℃ in a humidified 5% CO2 incubator. Cells were harvested from subcultures, supplemented with 0.01% trypsin EDTA (Sigma-Aldrich, St. Louis, MO, USA), and re-suspended in fresh medium for experiments.
siRNA and Doxorubicin
Survivin-targeted siRNA (Human BIRC5 [Gene ID 332], target sequence: CAAAGGAAACCAACAAUAA, GCAAAGGAAACCAACAAUA, CACCGCAUCUCUACAUUCA, CCACUGAGAACGAGCCAGA) was obtained from the siGENOME SMARTpool (Dharmacon Products, Thermo Fisher Scientific, Waltham, MA, USA), and adjusted according to the manufacturer's instruction. Doxorubicin (molecular weight, 579.98 g/mol) was purchased from Sigma-Aldrich, St. Louis, MO, USA.
MLC Preparation
Microbubble-liposome complexes were prepared as previously described (7). Lipid stocks (Avanti Polar Lipids, Albaster, AL, USA) comprising 15.4 mg of 1,2-dipalmitory-sn-glycero-3-phosphatidylcholine, 3.5 mg of cholesterol, 1 mg of dicetyl phosphate, 1.2 mg of 1,2-dipalmitory-sn-glycero-3-phosphoethanolamine, and 5 mg of 1,2-distearoyl-sn-glycero-3-phosphoethanolamine-N-(PDP[polyethylene glycol]-2000) were dissolved in 5 mL of 99.9% chloroform (Sigma-Aldrich). The mixture was lyophilized for 24 hours to remove chloroform, and mixed with 2 mL of solvent comprising glycerin, propylene glycol, and H2O (1:2:7). Cores of synthesized microbubbles were filled with sulfur hexafluoride gas (SF6). Liposomes were formed by freezing and thawing the mixture 5 times using liquid nitrogen, followed by agitation in a sonicator at 60℃ for 5 minutes with 2 mL of H2O. The resultant multilamellar vesicles were extruded using polycarbonate filters (filter size, 200 nm) to obtain liposomes smaller than 200 nm. Microbubbles and liposomes were shaken together at 25℃ for 2 hours to form MLC. Sulfosuccinimidyl-4-(N-maleimidomethyl) cyclohexane-1-carboxylate (5 mg; Sigma-Aldrich) was added to MLCs, followed by conjugation with anti-Her2 antibodies (Herceptin®; Roche, Berlin, Germany) by shaking at 4℃ for 24 hours to allow MLCs to target Her2-expressing prostate cancer cells (12).
Ultrasound-Mediated Intracellular Delivery of MLC
Microbubble-liposome complexes were labeled using fluorescein isothiocynate (Sigma-Aldrich) for microbubbles to emit green fluorescence, and Texas red (Sigma-Aldrich) for liposomes to emit red fluorescence. PC-3 and LNCaP cells were seeded in 1-well chamber slides (1 × 105 cells/well). The cells were subsequently mixed with MLCs and incubated for 3 hours at 37℃.
After washes with cold phosphate buffered saline (PBS), the cells were exposed to ultrasound waves using an ultrasound scanner (Philips Medical Systems, Bothell, WA, USA) with a linear probe. The cells were seeded in a line in the chamber slide, and submerged in buffered saline. Ultrasound exposure ("US-flashing") was performed by applying ultrasound waves ranging in frequency from 5 to 12 MHz over the cell chambers, with an interval of 1 second for 2 minutes and a mechanical index of 0.61.
Transfected cells were incubated for 3 hours at 37℃, and the intracellular MLCs were visually localized using a confocal laser scanning microscopy at × 400 magnifications (Leica Microsystems, Wetzler, Germany).
Loading of siRNA and Doxorubicin into MLCs
Microbubble-liposome complexes were loaded with survivin-targeted siRNA and/or doxorubicin in 4 formulations: 1) unloaded MLCs, 2) MLCs with doxorubicin (Dox-MLCs), 3) MLCs with siRNA (siRNA-MLCs), and 4) MLCs with siRNA and doxorubicin (Dox-siRNA-MLCs) by adding 5 nmoL of siRNA and/or 1 mg of doxorubicin to MLCs and dissolving the combination in 2 mL of H2O. After freezing, thawing, and agitation, final complexes were filtered using 200-nm polycarbonate filters.
Before doxorubicin loading, ultraviolet (UV) absorbance standard curve was obtained using a UV-visible (UV-vis) spectrophotometer (Scinco, Seoul, Korea). After loading, Dox-MLCs were separated from the free drug by centrifugation at 13000 rpm, with the resultant supernatant analyzed by UV-vis spectrophotometry. UV detection was maximal at 480 nm. The loading efficiency was calculated using the following equation: loading efficiency ([initial drug concentration - drug concentration in the supernatant] / initial drug concentration) × 100%.
Effect of Dox-siRNA-MLCs Delivery and Ultrasound Exposure on Cell Viability
PC-3 and LNCaP cells were seeded in a 96-well chamber slide at 2.0 × 103 cells/well. The cells were treated with 4 different methods: 1) no treatment (group 1), 2) Dox-siRNA-MLCs loading (group 2), 3) US-flashing without MLCs (group 3), and 4) Dox-siRNA-MLCs loading with US-flashing (group 4). For groups 2 and 4, Dox-siRNA-MLCs were added to each well, and the cells were incubated for 3 hours at 37℃. The cells in group 3 and 4 underwent US-flashing using the parameters described earlier.
Clonogenic MTT assay was performed immediately following treatment (day 0) and after 3 days of incubation (day 3). MTT reagent (50 µL; Sigma-Aldrich) was added to the treated cells, followed by 150 µL of DMSO (Sigma-Aldrich) and incubated for 3 hours at 37℃. Cell plates were shaken for 1 hour to dissolve MTT crystals. Optical density at 450 nm was read in a spectrophotometer (Scinco, Seoul, Korea), and cell viability was calculated relative to the control group.
Effect of Therapeutic Agents and Ultrasound Exposure on Cell Viability
The effect of each therapeutic agent (survivin-siRNA and doxorubicin) with or without ultrasound exposure was assessed by MTT assay. PC-3 and LNCaP cells (2.0 × 103 cells/well) were seeded in 96-well chambers. The cells were treated with 1) no additives, 2) unloaded MLCs, 3) Dox-MLCs, 4) siRNA-MLCs, or 5) Dox-siRNA-MLCs. A separate group of cells underwent same treatment, with the addition of US-flashing performed as previously described. After 3 days of incubation at 37℃, viability of treated cells was evaluated using the MTT assay.
In Vivo Assays
All animal protocols were approved by the Institutional Animal Care and Use Committee. PC-3 and LNCaP cells (1.5 × 106 cells in 0.2 mL of PBS) were subcutaneously injected in both flanks of 6 athymic nude male mice (n = 3 for each cell type) from an animal facility (Orient, Seoul, Korea) to produce xenografts of prostate tumor model.
After 4 to 6 weeks of tumor growth, mice were euthanized with isoflurane. One mouse with a PC-3 tumor and one with an LNCaP tumor were used as controls without any treatment. Two mice in each group were injected 0.2 mL of Dox-siRNA-MLC dissolved in PBS via tail vein. All MLCs were fluorescence-labeled with Texas red. Following the injection, US-flashing was performed for 5 minutes with an interval of 3 seconds with the mechanical index of 0.47 on the tumors in the right flank (Fig. 1). US-flashing was not applied to the left flank tumor, to allow the two tumors to be compared within the same animal.
After 24 hours, mice were sacrificed and tissue sections were obtained from tumors on each side of the animal. Tumor uptake of Dox-siRNA-MLC was assessed by confocal laser scanning microscopy at × 400 magnifications and survivin expression was quantified by Western blot analysis.
Western Blot Analysis
Tissue samples were homogenized in 600 µL of PROPREPTM Protein Extraction solution (Intron Biotechnology, Seoul, Korea). After centrifugation at 13000 rpm for 10 minutes at 4℃, 20 µg of supernatant was added to a 5 × SDS gel-loading buffer. The sample solution was boiled at 100℃ for 5 minutes, loaded onto the SDS gel, and electrophoresis was performed for 20 minutes at 80 V and 60 minutes at 130 V. Proteins were transferred to a membrane in transfer buffer at 80 V for 1.5 hours. The membrane was blocked with 5% skim milk in Tris-buffered saline with Tween (TBS-T) solution for 30 minutes at room temperature, and incubated with a diluted solution of primary antibody (anti-survivin, 1:2000 dilution; β-actin, 1:10000 dilution) overnight at 4℃. Following washing in TBS-T, the membrane was incubated with secondary antibody solution (anti-rabbit, 1:2000 dilution) for 1 hour at room temperature. Proteins of interest were detected using WEST-ZOL® Western Blot Detection System (Intron Biotechnology, Seoul, Korea). Survivin expression was normalized to β-actin levels, and the ratio of survivin expression relative to β-actin was calculated.
Statistical Analysis
Data were expressed as means ± standard deviations. Differences between multiple experimental groups were compared using Kruskal-Wallis tests followed by post-hoc tests with Bonferroni correction. Comparisons between two experimental groups were performed with Mann-Whitney or Wilcoxon signed rank tests. Statistical analyses were performed using statistical software (SPSS, version 18.0; SPSS Inc., Chicago, IL, USA). p values < 0.05 were considered statistically significant.
RESULTS
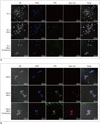
Efficiency of Target-Specific Intracellular Uptake
No substantial fluorescence was observed before and after US-flashing in PC-3 cells that have relatively low Her2 expression (Fig. 2A). Conversely, LNCaP cells, which are known to express higher levels of Her2 than PC-3 cells, showed substantial green and red fluorescence, indicating the presence of labeled microbubbles and liposomes after incubation with the mix of MLCs, both before and after US-flashing (Fig. 2B).
Efficiency of Doxorubicin Loading in Synthesis of MLC
The efficiency of doxorubicin loading was determined as 61.9%, with the total concentration of loaded doxorubicin of 213.6 µM. The concentration of loaded doxorubicin per treated cell well was 21.4 µM.
Effect of Dox-siRNA-MLCs Delivery and Ultrasound Exposure on Cell Viability
Figure 3A summarized the cell survival data acquired following different treatments.
In PC-3 cells (Fig. 3B), cell survival rate was determined as > 90% in all treatment groups on Day 0. While cell survival rate was reduced by 4% in group 4 on Day 3, no statistically significant difference from Day 0 was observed (Wilcoxon signed rank test, p = 0.25). The other 3 groups also did not show any significant alterations in cell viability on Day 3 (Wilcoxon signed rank test, group 1, p = 0.73, group 2, p = 0.46, group 3, p = 0.05).
In LNCaP cells (Fig. 3C), cell viability on Day 0 was > 90% in all treatment groups. After 3 days of incubation, group 4 (cells treated with Dox-siRNA-MLCs and US-flashing) showed a significant reduction in cell viability (94.2 ± 2.8%, vs. 41.8 ± 3.2%, on Days 0 and 3, respectively; Wilcoxon signed rank test, p = 0.009). Cell viability was significantly different between the 4 treatment groups on day 3 (Kruskal-Wallis test, p = 0.006). In subgroup analysis, cell viability of group 4 was significantly lower than in group 1 (Mann-Whitney U test, p = 0.005).
Effect of Therapeutic Agents and Ultrasound on Cell Viability
Figure 4A summarized the cell viability data in subgroups
of PC-3 and LNCaP cells.
Cell survival rate in PC-3 cells was > 90% in all subgroups without US-flashing (Fig. 4B), which was not significantly altered in cells treated with additional US-flashing (all subgroups, Mann-Whitney U test, p > 0.05).
Viabilities in LNCaP cells treated with Dox-MLCs, siRNA-MLCs, or Dox-siRNA-MLCs were significantly lower in groups with US-flashing (Dox-MLCs, 88.0 ± 3.4% vs. 63.0 ± 1.8%, p = 0.009; siRNA-MLCs, 87.0 ± 4.1% vs. 73.0 ± 3.8%, p = 0.009; Dox-siRNA-MLCs, 85.0 ± 2.9% vs. 55.0 ± 3.5%, p = 0.009, Mann-Whitney U test). The rate of reduction of cell viability was highest in cells treated with Dox-siRNA-MLCs and US-flashing, followed by the cells treated with Dox-MLCs and US-flashing, and siRNA-MLCs and US-flashing (Fig. 4C). The viability in cells treated with Dox-siRNA-MLCs and US-flashing was significantly lower than in cells treated with Dox-MLCs and US-flashing (Mann-Whitney U test, p = 0.008).
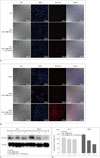
Suppression of Survivin Expression after Treatment with Dox-siRNA-MLC and Ultrasound
In PC-3 xenograft tumor model, no substantial uptake of Dox-siRNA-MLCs in the tumor was observed by confocal microscopy (Fig. 5A). In contrast, substantial red fluorescence reflecting tumor uptake of Dox-siRNA-MLCs was observed in LNCaP tumor, which became more pronounced after US-flashing (Fig. 5B).
In Western blot analysis, the levels of survivin expression were lower in LNCaP tumors treated with Dox-siRNA-MLC compared to the control, and lowest in LNCaP tumors treated with both Dox-siRNA-MLC and US-flashing. Survivin expression ratios were 77.4 ± 4.90% in control tissues, 52.7 ± 2.83% in LNCaP tumors with Dox-siRNA-MLCs, and 36.7 ± 1.34% in LNCaP tumors with Dox-siRNA-MLCs and US-flashing (Kruskal-Wallis test, p = 0.027; all subgroups, Mann-Whitney U test, p = 0.10) (Fig. 5C, D). Survivin expression in the treated PC-3 tumors was decreased to a lesser degree (control, 63.1 ± 4.36%; PC-3 tumor with Dox-siRNA-MLCs, 56.8 ± 4.35%; PC-3 tumor with Dox-siRNA-MLCs and US-flashing, 56.6 ± 3.08%; Kruskal-Wallis test, p = 0.113; subgroup analysis, Mann-Whitney U test, p = 0.20, 1.0, 0.10, respectively).
DISCUSSION
Figure 6 depicted schematically ultrasound and MLC-mediated delivery of survivin-targeted siRNA and doxorubicin into prostate cancer cells. We demonstrated that MLCs could effectively target prostate cancer cells expressing Her2 by the conjugation of anti-Her2 antibodies. We successfully loaded MLCs with survivin-targeted siRNA and doxorubicin, and delivered into prostate cancer cells in vitro and prostate tumor model in vivo. Additional ultrasound exposure escalated the therapeutic effects of the loaded substances.
Most prostate cancers tend to recur as aggressive androgen-refractory prostate cancers (1314). While chemotherapeutic drugs have improved the survival rate (15), no reliable therapeutic method for androgen-refractory prostate cancer has yet been established. In this regard, siRNA in prostate cancer therapy has been actively investigated over the years. Survivin, an anti-apoptotic protein (1617) strongly expressed in prostate cancer cells (18), has been identified as an attractive therapeutic target. It is associated with cellular proliferation (19), resistance to androgen deprivation therapy, and metastasis of prostate cancer (1920). The siRNA-mediated down-regulation of survivin can reduce prostate cancer cell proliferation, stimulate apoptosis (1617), and increase the chemosensitivity (2122).
However, intracellular delivery of siRNA is hindered by its fast degradation in the physiologic environment and its inability to cross the cell membrane (45). Additive "carriers" must therefore be attached to siRNA to achieve effective delivery. These carriers need to protect siRNA from degradation, transport it to the target cells, and release it into the cytoplasm without toxicity (46). MLCs contain microbubbles that function effectively as carriers, and can target specific tissues by incorporating target-specific ligands (8). Plus, ultrasound can facilitate the delivery of molecules by sonoporation (89). Additional disruption of microbubbles by ultrasound energy can increase cell permeability (23), while tissue-selective targeting also can be achieved by focusing ultrasonic field (10). Thus, ultrasound MLC-mediated delivery can be a potent tool for target-specific therapy in prostate cancer.
One novel finding of our study was from the attachment of anti-Her2 antibodies to MLCs. The cellular level approach of the cancer treatment necessitates proper targeting of cancer cells. Thus, specific targeting method suitable for prostate cancer cells is required. Since it is well known that prostate cancer cells express Her2 (12), we aimed to facilitate the ability of MLCs to target prostate cancer cells by conjugating anti-Her2 antibodies. As a result, intracellular uptake of MLCs was significantly higher in LNCaP cells that express higher levels of Her2 than PC-3 cells. This proved that MLCs conjugated with anti-Her2 antibodies could effectively target prostate cancer cells.
The cytotoxic effect was accentuated with the delivery of MLCs containing both doxorubicin and survivin-targeted siRNA, as compared to the MLCs containing doxorubicin alone. Knock-down of survivin was previously shown to increase the chemosensitivity of prostate cancer (2122). Therefore, co-delivery of chemotherapeutic agent with siRNA can elicit a synergistic effect in prostate cancer treatment. Since MLCs can deliver both siRNA and doxorubicin, it can be an effective tool for targeted cancer therapy.
Targeting of MLCs and their capacity to deliver siRNA and doxorubicin was verified in a xenograft prostate tumor model as well. Suppression of survivin expression was observed in the tumor tissue, especially with the application of ultrasound waves. This result suggested that the delivery of siRNA and doxorubicin could be improved by ultrasound-induced microbubble burst, both in vitro and in vivo.
There are a few limitations in our study. First, fluorescent imaging was performed 3 hours following the start of incubation, which could result in an underestimation of the total MLC uptake, since the delayed uptake would be excluded from our analysis. Further studies need to analyze the delayed uptake following sonoporation to support our findings. Second, a small number of mice were included in vivo analysis, precluding any statistical analysis. While our observations demonstrate the feasibility of in vivo application of ultrasound MLC-mediated delivery of therapeutic agents, further studies are needed to validate our results.
In conclusion, MLCs loaded with specific ligand are an effective tool for intracellular delivery of siRNA and doxorubicin into prostate cancer cells in vitro and into a prostate tumor model in vivo. With the application of ultrasound, MLCs containing survivin-targeted siRNA and doxorubicin elicited a cytotoxic effect and inhibited the expression of survivin. Therefore, ultrasound MLC-mediated delivery of siRNA and chemotherapeutic agents opens up the possibility of clinically applicable image-guided therapy and provides novel prospects for therapeutic applications in the near future.




 XML Download
XML Download