

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
The survival of several types of secretary epithelia is affected by fluid and HCO3− secretion. HCO3− regulates the pH of human body fluids and accelerates solubilization of macromolecules and ions in secreted fluids. The failure of this mechanism could result in the partial damage of tissues that affects the pancreas, lung epithelium, and vas deferens in cystic fibrosis (CF). Therefore, knowledge of fluid and HCO3− secretion in CF could contribute to the discovery of new methods that end the progress of the disease. The pancreatic duct specializes in fluid and HCO3− secretion. Hence, we use this as a perfect model to study this major function of secretory epithelia. The secretion of HCO3− rich fluid occurs in the pancreas. The synthesis and secretion of the digestive enzymes occurs in acini, and the secretion of NaCl-rich isotonic fluid occurs in order to cleanse digestive enzymes [1]. The fluids and HCO3− are secreted via the pancreatic duct and then the Cl− is absorbed to create pancreatic juice, which includes approximately 140 mM of HCO3− and 20 mM of Cl− in humans [2].
Pancreatic Na+/HCO3− cotransporter (pNBC1) is the Na+/HCO3− cotransporter isoform of the pancreatic ductal cells originally cloned from human pancreas using polymerase chain reaction [3]. The name pNBC1 was recently changed to NBCe1-B based on an electrogenic NBCe1 subgroup of the Na+-coupled HCO3− transporter (NCBT) supergroup [45], which exists on the basolateral membrane (BLM). We found strong NBC activity in the pancreatic duct of Rattus norvegicus [6]. Similarly, this was also reported in the pancreatic ducts of Cavia porcellus [78]. The electrogenic transporter, NBCe1-B, exhibits a 1:2 Na+ to HCO3− stoichiometry in pancreatic ductal cells [9]. Even if the Na+/HCO3− cotransport stoichiometry does not occur in the duct, NBCe1-B is involved in HCO3− inflow via the stimulated duct, BLM [10], which also requires a 1:2 Na+ to HCO3− stoichiometry. NBCe1-B activity is significantly downregulated by with-no-lysine kinase (WNK) and Ste20-related proline-alanine-rich kinase (SPAK) [11], and upregulated by inositol 1,4,5-triphosphate (IP3) receptors binding protein released with IP3 (IRBIT) [11121314]. The recruitment of PP1 to IRBIT facilitates the activation of NBCe1-B. This recruitment also reduces the inhibitory WNK/SPAK effect via NBCe1-B dephosphorylation [11].
PP1 is a dual-specific protein phosphatase that mediates various cellular systems such as neuronal signaling, transcription, cell cycle progression, carbohydrate metabolism, muscle contraction, and protein synthesis [15161718]. Additionally, PP1 is reported to control the activity and/or surface distribution of many ion channels, receptors, and transporters [1920212223]. PP1 also controls the dephosphorylation of many cellular target proteins by associating multiple binding proteins with PP1c. This interaction uses two kinds of PP1-binding consensus residues—FxxR/KxR/K and R/Kx(0,1)VxF—as specific codes so as to recognize proteins as binding partners of PP1c [24].
In the current study, we probed PP1 involved in the activation of NBCe1-B. The results showed that NBCe1-B included the specific PP1-binding consensus residues, FxxR/KxR/K. By performing competition experiments and site-directed mutagenesis, we found that this binding site played a major role in PP1's interaction with NBCe1-B in vitro. Therefore, we strongly concluded NBCe1-B to be an important PP1-binding partner. We have also attempted to elucidate the differences in the regulatory mechanisms between the physiological functions of the NCBT superfamily via PP1.
Go to : 
METHODS
Solutions, materials, mutagenesis, and plasmid construction
The standard bath solution was made using 10 mM of HEPES (pH 7.4), 140 mM of NaCl, 1 mM of MgCl2, 5 mM of KCl, 1 mM of CaCl2, and 10 mM of glucose. HCO3−-buffered solution consisted of substitution Na+-HCO3− (25 mM) for Na+ (25 mM) salts and by reduction the HEPES concentration (2.5 mM) and was gassed using CO2 (5%) and O2 (95%). Na+-free solution consisted of substitution N-Methyl-D-glucamine for Na+. The osmolarity of every solution was set at 310 mOsm. Co-immunoprecipitation assay antibodies for PP1 and GFP were purchased from Santa Cruz and Invitrogen, respectively. Point and deletion mutants were made via site-directed mutagenesis using the QuikChange® Site-Directed Mutagenesis Kit (Stratagene). PP1-binding consensus residue deletion mutant (deletion of 922FMDRLK927 amino acids) and 3-point mutant (F922A, R925A, and K927A) were made via PCR splicing with template and point mutagenesis, respectively. NBCe1-B and PP1 constructs were designed based on previous studies [1114]. The NBCe1-B gene was inserted into the pCEGFP-C1 vector (Clontech). PCR was performed using the following designated primers to substitute 3 amino acids and delete the PP1-binding consensus residue region: 5′-CTT AAT GGT GTG CAG GCC ATG GAT GCT CTG GCG CTG CTT CTG ATG CCT-3′ (3-point mutant); 5′-TCC CTT AAT GGT GTG CAG CTG CTT CTG ATG CCT CTG-3′ (PP1-binding consensus residue deletion mutant). Every construct was verified via sequence analysis.
Cell transfection
All transfections in HEK293T and HeLa cells were carried out using Lipofectamine® 3,000 Transfection Reagent (Invitrogen). To obtain high transfection efficiency, we used HEK293T cells for biotinylation and co-immunoprecipitation assays. For pHi measurements, we required the cells to tolerate fast flow rates. Thus, we choose HeLa cells for their ability to attach strongly onto the coverslips. Preliminary experiments were performed to determine the optimal transfection conditions. When cells were transfected with multiple plasmids, the transfection ratio was set in the preliminary experiments and the total number of plasmids were kept constant for all conditions by using empty vectors. Transfected cells were identified and co-transfected with the GFP plasmid. GFP fluorescence intensity was used to determine cells showing similar expression level as co-transfected plasmids.
Measurement of intracellular pH
Intracellular pH (pHi) was measured by recording the fluorescence ratio of BCECF (TEFlabs) at 490 and 440 nm excitation wavelengths and by calibrating the fluorescence signals as indicated previously [14]. The fluorescence ratio was recorded using clusters of 2–5 transfected HeLa cells [14]. HeLa cells were loaded with BCECF for a 20 min incubation at room temperature, and then perfused with HEPES-buffered medium for at least 5 min before measuring pHi. Na+-HCO3− cotransport was induced via acid loading by perfusing the cells with Na+-free HCO3−-buffered medium containing 10 µM of S-(N-ethyl-N-isopropyl) amiloride, which blocks the Na+/H+ exchanger (NHE) [25]. NBCe1-B activity was initiated by perfusing the HeLa cells with HCO3−-buffered media containing 140 mM of Na+. The rates of Na+-dependent changes in pHi (Δ pHi (recovery)/Δ pHi (total)) demonstrated NBCe1-B activity. Moreover, NBCe1-B activity was also estimated from the slopes of the first derivatives of pHi increase in HCO3−-buffered media containing 140 mM of Na+, labeled as Δ pHi/min.
Co-immunoprecipitation and biotinylation
For co-immunoprecipitation assays, transfected HEK293T cell extracts were prepared by disrupting the cells in ice-cold lysis buffer (20 mM Tris, 150 mM NaCl, 2 mM EDTA, 1% Triton X-100, and protease inhibitor cocktail), and incubating them for 15–30 min followed by centrifugation at 20,000 ×g for 30 min. After protein quantification using the DC Protein Assay (Bio-Rad), the extracts containing 500 µg of protein were then incubated with 1 µg of anti-GFP or anti-HA antibody beads. The antibody beads were then collected and washed thrice with lysis buffer. Proteins bound to the beads were recovered after incubating the beads in 25 µL of SDS sample buffer with dithiothreitol (DTT) at 60℃ for 30 min. The extracts, 1% of the input (3 µg) and 80% of the immunoprecipitates (20 µL), were loaded and separated by 10% SDS-PAGE, transferred to PVDF membranes (Bio-Rad), and detected with HRP-conjugated anti-GFP or anti-PP1 antibodies (Santa Cruz).
To verify NBCe1-B surface expression via biotinylation assay, transfected HEK293T cells were washed with 100 mM PBS/glycine before subjecting to 0.1% Sulfo-NHS-LC-Biotin (Abcam) for 2 h in a cold room, followed by lysis with 1% PBS/Triton X-100. The lysate was mixed with avidin beads and incubated for 2 h in a cold room. The avidin/biotin bead complex was washed thrice with 1% PBS/Triton X-100. Biotinylated NBCe1-B was eluted in SDS sample buffer with dithiothreitol (DTT). We used the ImageJ program (NIH, Bethesda, MD, USA) to quantify and determine the intensity of the blot.
Densitometric quantification
To calculate the abundant binding of NBCe1-B with PP1 or surface expression of NBCe1-B indicated as the amount of NBCe1-B co-precipitates or surface expression divided by the total amount of the NBCe1-B input as follows: NBCe1-BBlot/NBCe1-BInput (for comparison, every result was normalized to 100% for NBCe1-B). *p<0.05 vs. wild type (n=3).
Statistical analysis
Results of all experiments are indicated as mean±SEM for the number of indicated experiments. Significant differences (p<0.05) between the wild type and each experimental group were statistically determined by one-way ANOVA.
Go to :
RESULTS
NBCe1-B contains PP1-binding consensus residues
We performed multiple alignment analysis of the C-terminal part of the human NBCe1-B (Q9Y6R1.1) against the orthologous transporters of Mus musculus (O88343.2), Rattus norvegicus (Q9JI66.1), Bos taurus (NP_777030.1), and Trichosurus vulpecula (AEQ33587.1). This result showed that residues 910-960 were well conserved between the organisms, except for residue 955 in Mus musculus (Fig. 1A). A sequence search revealed that NBCe1-B contained the PP1-binding consensus residues, 922FMDRLK927, which were required for the PP1-binding consensus residues, FxxR/KxR/K [24]. This sequence is conserved from Homo sapiens to Trichosurus vulpecula (Fig. 1A).
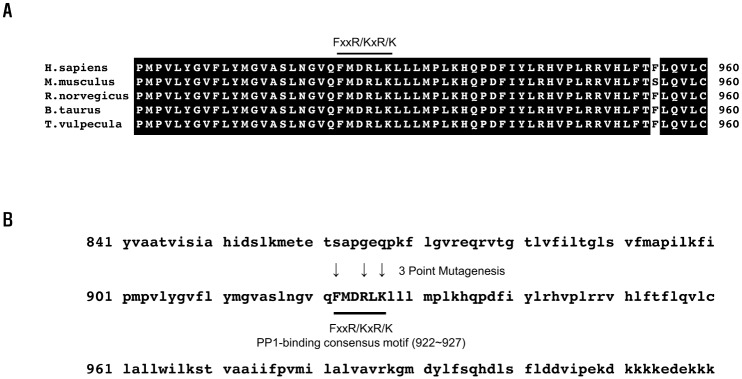 | Fig. 1Conservation of the PP1-binding consensus residues in NBCe1-B.(A) Sequence alignment analysis: the C-terminal part of the human NBCe1-B (Q9Y6R1.1) against the orthologous transporters of Mus musculus (O88343.2), Rattus norvegicus (Q9JI66.1), Bos taurus (NP_777030.1), and Trichosurus vulpecula (AEQ33587.1). (B) Amino acids (downward-pointing arrows) are critical in binding to PP1, and substituted by site-directed mutagenesis. (A, B) The PP1-binding consensus residues are indicated using bold bars.
|
To verify the applicability of the FxxR/KxR/K residues of NBCe1-B as a PP1-binding consensus residue, we constructed the 3-point mutant (F922A, R925A, and K927A) and the PP1-binding consensus residue deletion mutant (deletion of amino acids 922FMDRLK927). Substitution of F922, R925, and K927 with alanine or deletion of the 922FMDRLK927 residues was achieved via site-specific mutagenesis (Fig. 1B).
PP1 directly interacts with PP1-binding consensus residues of NBCe1-B in vitro
We previously reported that IRBIT recruited PP1 to NBCe1-B [11]. However, the PP1-binding consensus residues in NBCe1-B have not been studied yet. To determine that the 922FMDRLK927 residues in NBCe1-B were critical for its interaction with PP1 in vitro, co-immunoprecipitation assay was performed between NBCe1-B and the 2 mutants i.e., the alanine-replaced F922, R925, and K927 mutant and the PP1-binding consensus residue deletion mutant. As shown in Fig. 2A, wild type NBCe1-B strongly interacted with PP1. However, the alanine-substituted F922, R925, and K927 residues and the deleted 922FMDRLK927 residues of NBCe1-B did not bind to PP1 (Fig. 2A), significantly reducing the interaction with PP1 to 28.9±9.3% and 18.2±9.1% (Fig. 2B). This result confirms that the PP1-binding consensus residues, 922FMDRLK927, play an essential role in interacting with PP1.
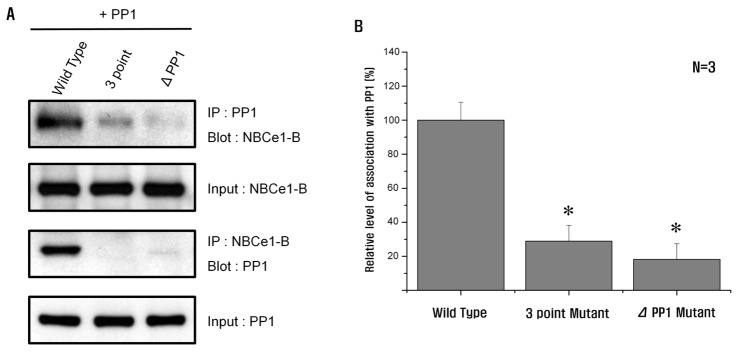 | Fig. 2PP1 interacts with the PP1-binding consensus residues in NBCe1-B.(A) Co-immunoprecipitation of NBCe1-B with PP1 in HEK293T cells transfected with PP1 co-expressing NBCe1-B, 3-point mutant, and PP1-binding consensus residue deletion mutant. The cells were lysed with RIPA buffer. The extract was immunoprecipitated with anti-GFP or anti-PP1. (B) HEK293T cells transfected with 3-point mutant or PP1-binding consensus residue deletion mutant showing a significant decrease of NBCe1-B activity via association with PP1 (28.9±9.3% and 18.2±9.1%; n=3; *p<0.05). *Significance was compared against the wild type value.
|
NBCe1-B activity is controlled by PP1
We reported previously that, like IRBIT, overexpression of PP1 regulated NBCe1-B activity [11]. To identify the function of PP1 on the PP1-binding consensus residues in NBCe1-B, we recorded the change of NBCe1-B activity in each designed condition by measuring the pHi recovery from acidification. Thus, we determined NBCe1-B activity using the pH indicator, BCECF-AM (2′,7′-bis(2-carboxyethyl)-5(6)-carboxyfluorescein tetrakis(acetoxymethyl) ester), to measure pHi. To eliminate any unnecessary contribution to pHi recovery, we used an anion exchange inhibitor, DIDS (disodium 4,4′-diisothiocyanatostilbene-2,2′-disulfonate), and an NHE (Na+/H+ exchanger) inhibitor, EIPA (5-(N-ethyl-N-isopropyl)-amiloride) [14]. By mutating the PP1-binding consensus residues, the interaction with PP1 was inhibited, which further inhibited NBCe1-B activity (Figs. 3B and C). This result suggested that the interaction between NBCe1-B and PP1 could mediate cell surface localization and activity of the transporter. Therefore, the recruitment of PP1 to NBCe1-B prevented the inhibitory SPAK effect toward NBCe1-B activation.
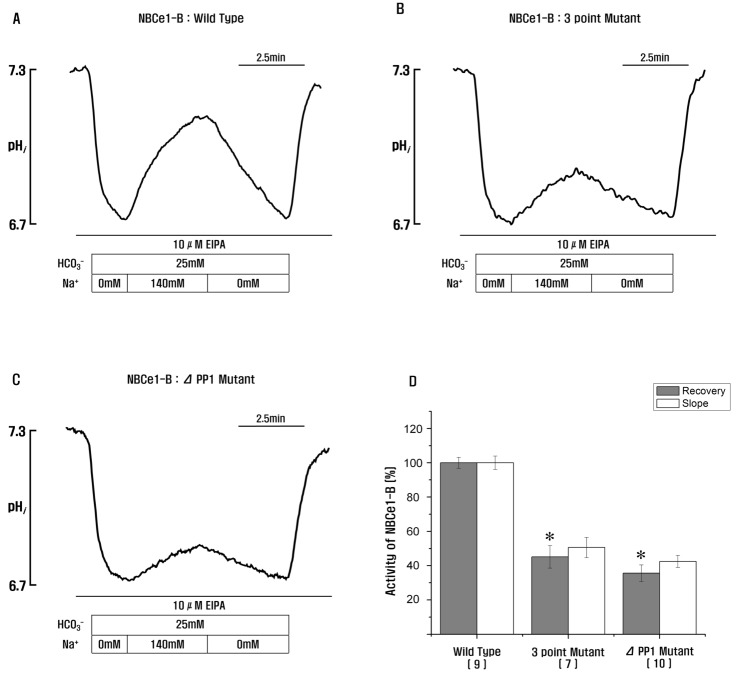 | Fig. 3Role of PP1-binding consensus residues in NBCe1-B.(A–C) Measurement of NBCe1-B activity in HeLa cells transfected with wild type, 3-point mutant, and PP1-binding consensus residue deletion mutant by monitoring intracellular pH. (D) HeLa cells transfected with 3-point mutant or PP1-binding consensus residue deletion mutant showing a significant decrease of NBCe1-B activity (45.1±6.5% and 35.7±4.7%; *p<0.05). *Significance was compared against the wild type value. Number of experiments for each condition are shown below the X-axis. Recovery, the rates of Na+-dependent changes in pHi (Δ pHi (recovery)/Δ pHi (total)); Slope, the slopes of the first derivatives of pHi increase in HCO3−-buffered media containing 140 mM of Na+(Δ pHi/min).
|
Next, we recorded the pHi difference in HeLa cells co-expressing NBCe1-B mutants with PP1 to test the function of PP1 in NBCe1-B activation. The overexpression of PP1 stimulated NBCe1-B activity as in the previous result (Fig. 4B). More importantly, PP1 could still not activate NBCe1-B inhibited either by the 3-point mutant (Fig. 4C) or by the PP1-binding consensus residue deletion mutant (Fig. 4D), revealing the PP1-dependent activity of NBCe1-B. PP1 consistently and specifically bound to a part of the NBCe1-B C-terminal and mediated the activation of NBCe1-B.
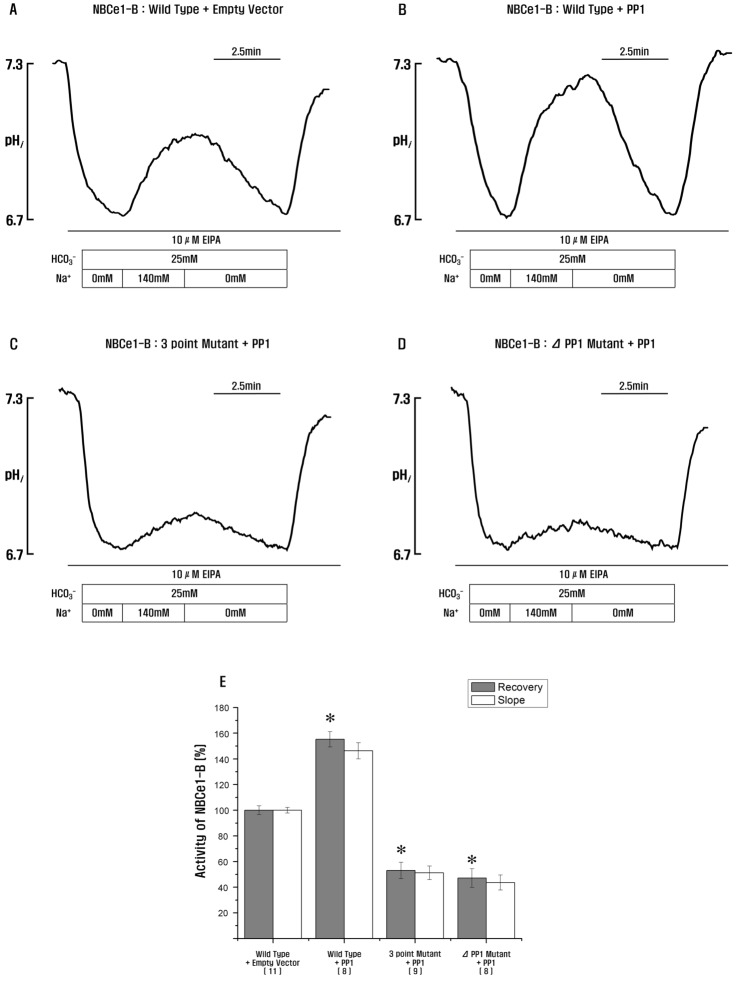 | Fig. 4PP1 stimulates NBCe1-B activity via PP1-binding consensus residues in NBCe1-B.(A–D) Measurement of NBCe1-B activity in HeLa cells transfected with wild type, 3-point mutant, and PP1-binding consensus residue deletion mutant co-expressing PP1 by monitoring intracellular pH. (E) NBCe1-B with PP1 shows a significant increase in NBCe1-B activity, but 3-point mutant and PP1-binding consensus residue deletion mutant co-expressing PP1 inhibit NBCe1-B activity (155.3±5.9%, 53.1±6.4% and 47.2±7.3%; *p<0.05). *Significance was compared against the wild type+empty vector value. Number of experiments for each condition are shown below the X-axis. Recovery, the rates of Na+-dependent changes in pHi (Δ pHi (recovery)/Δ pHi (total)); Slope, the slopes of the first derivatives of pHi increase in HCO3−-buffered media containing 140 mM of Na+(Δ pHi/min).
|
According to the results, we demonstrated that PP1 was associated with HCO3− secretion after stimulation. The PP1-mediated dephosphorylation of NBCe1-B was likely related to the effect of PP1. However, PP1-mediated dephosphorylation of the other associated proteins such as SPAK, WNK, or scaffold proteins was certainly due to the effect of PP1. A recent study revealed that the inhibition of the stimulatory SPAK effect on the activation of NKCC1 was due to the dephosphorylation of SPAK via PP1 [26]. A similar mechanism may apply with respect to the effects of PP1 and inhibitory SPAK on the activation of NBCe1-B.
PP1 regulates surface expression of NBCe1-B
A biotinylation assay was performed to identify and measure the effects of PP1 on NBCe1-B cell membrane expression levels in HEK293T cells. First, we confirmed that the wild type, 3-point mutant, and PP1-binding consensus residue deletion mutant NBCe1-B surface expression level was determined via blotting with anti-GFP (Fig. 5A). The expression levels of the 3-point mutant and PP1-binding consensus residue deletion mutant showed a 5-fold decrease in NBCe1-B expression on the cell surface when compared to the NBCe1-B expression levels of the wild type on the cell surface (Fig. 5B). This result suggested that the PP1-binding consensus residues, 922FMDRLK927, were involved in the control of membrane expression. A previous study reported that PP1 increased the activity of NBCe1-B in vitro [11]. Based on this result, we found that PP1 increased NBCe1-B surface expression (Fig. 5A). Using the 3-point mutant and PP1-binding consensus residue deletion mutant, we found that F922, R925, and K927 were the major factors that controlled the NBCe1-B cell surface expression (Fig. 5A). Thus, we concluded that NBCe1-B and PP1 interaction increased the surface expression of NBCe1-B. Moreover, the PP1-mediated NBCe1-B dephosphorylation sufficiently increased NBCe1-B surface expression.
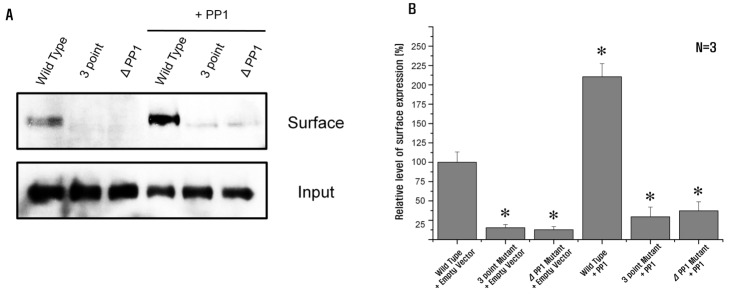 | Fig. 5NBCe1-B surface expression is associated with PP1.(A) Biotinylation of NBCe1-B with or without PP1 in HEK293T cells transfected with or without PP1 co-expressing NBCe1-B, 3-point mutant, and PP1-binding consensus residue deletion mutant. Cells were incubated with biotin. Biotinylated NBCe1-B was isolated and recovered. Input corresponds to 5% of the total NBCe1-B used for the avidin pull-down step. (B) NBCe1-B with PP1 shows an approximate 2-fold increase of NBCe1-B surface expression level (210.5±16.9%). However, 3-point mutant and PP1-binding consensus residue deletion mutant with or without PP1 significantly inhibit NBCe1-B surface expression level (29.5±12.6%, 37.2±11.8%, 15.3±4.2% and 12.6±4.3%, respectively). Significant differences (*p<0.05) between wild type and each experimental group were statistically determined by one-way ANOVA.
|
Go to :
DISCUSSION
PP1 is associated with the regulation of many ion transporters and channels. PP1 specifically interacts with RVXF residues existing in the NKCC1 N-terminal, and prevents the WNK/SPAK pathway from increasing NKCC1 functionality [26]. Moreover, PP1 also regulates the tetrameric ClC-2 channel when PP1 isoforms bind to the ClC-2 channel. However, calyculin A, a PP1/PP2A inhibitor, reduces the ClC-2 channel currents [22].
More importantly, PP1 was reported to be a key phosphatase that dephosphorylated the serine/threonine residues that were phosphorylated via the WNK/SPAK pathway. Some ion transporters and channels consist of PP1-binding consensus residues and a search analysis for these sequences revealed the existence of such residues in ROMK1, KCC, NHE3, and NBCe1-B [27]. Additionally, PP1-binding consensus residues may also be found in WNK1 and WNK4 [28]. Furthermore, PP1 is associated with regulating the activities of NKCC1 [2629], KCC [30], NBCe1-B, and CFTR [11]. In a previous study, we determined that IRBIT interfered with PP1's effects [11]. It was reported that IRBIT consisted of PP1-binding consensus residues [31]. PP1 was recruited to NBCe1-B by IRBIT, and this process was essential in inverting the WNK/SPAK pathway's effects and regulating transporters [11]. Tautomycin (the pharmacological inhibitor) or I2 (the native PP1 inhibitor) inhibited cotransporters by inhibiting PP1 and functioned like WNKs and SPAK [11]. Moreover, mutation of the PP1-binding consensus residues in IRBIT inhibited the interaction of PP1 with CFTR and NBCe1-B [11].
To date, the regulation of the cotransporters via the IRBIT/PP1 and WNK/SPAK pathways is considered to be a major regulatory pathway in many secretary epithelia. Recent studies revealed that WNKs were identified in a search for homologs of MAPK/ERK [32]. Studies also revealed that hypertension was associated with mutations in WNKs [33]. Additionally, the interaction between SPAK/OSR1 and WNKs, and their role in general regulatory signal transduction [3435], CFTR control, NBCe1-B expression control via IRBIT [14], and PP1 and WNK/SPAK interaction [11] were also discovered. Moreover, the control system using the IRBIT/PP1 and WNK/SPAK signal transduction may also establish a general pathway that decides the stimulatory secretory and resting states.
However, the mechanism of action between PP1 and NBCe1-B still needs to be elucidated. Previously, we proved that PP1 dephosphorylated NBCe1-B via radiography experiments [11]. In this study, we identified that PP1, like IRBIT, specifically recognized and bound to the PP1-binding consensus residues in NBCe1-B and considerably increased its activity. These results suggested that NBCe1-B and PP1 were interaction partner proteins and that PP1 was a multiple regulator not only by controlling NBCe1-B activity, but also by assisting IRBIT functionality via direct binding and dephosphorylation. This finding also proved that one of the most important functions of PP1 was to regulate pHi via NBCe1-B and IRBIT. The maintenance of an appropriate pHi is of utmost importance to a cell's normal physiology. pHi changes associated with acute metabolic activity signals the rearrangement of cellular function. NBCe1-B transports HCO3− across the plasma membrane, regulates pHi and transepithelial movement of acid-base equivalents, and plays an important role in the acid-base regulation of many epithelial cells. Thus, measuring pHi serves as a suitable tool to discover the regulatory process and physiological role of NBCe1-B.
Mutations in the PP1-binding consensus residues of NBCe1-B results in the decreased surface expression of NBCe1-B, which could impair an important mechanism such as NBCe1-B-mediated endocytosis. We found that the interaction between NBCe1-B and PP1 formed a critical protein for IRBIT regulation of NBCe1-B activity. The diagram illustrating the NBCe1-B regulatory mechanism via the IRBIT/PP1 and the WNK/SPAK pathways is displayed in Fig. 6. In these results, the increased NBCe1-B endocytosis caused by the PP1-binding consensus residue mutations in NBCe1-B may be due to the enhanced effects of WNK/SPAK and intersectin (ITSN). Since ITSN interacts with WNKs, this binding effect is an important step in stimulating ROMK1 endocytosis via WNKs [36].
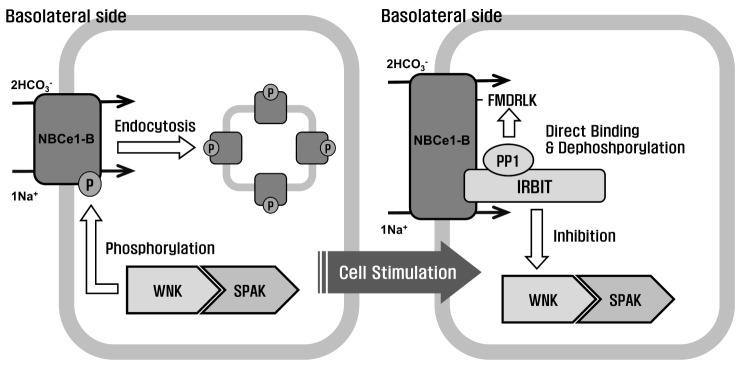 | Fig. 6Regulation of NBCe1-B via the IRBIT/PP1 and WNK/SPAK pathways.The WNK/SPAK pathway keeps the epithelial ductal cells in a steady state by decreasing the surface expression of NBCe1-B. The WNKs directly bind to SPAK, which phosphorylates NBCe1-B. After cell stimulation, IRBIT inhibits the WNK/SPAK pathway and activates NBCe1-B. IRBIT interacts with PP1 and PP1 is recruited to NBCe1-B by IRBIT. Thus, PP1 specifically docks with the PP1-binding consensus residue in NBCe1-B and dephosphorylates it, thereby restoring its surface expression.
|
However, PP1's association and stimulatory activity with respect to transporters still remains to be determined, except for its activity on NBCe1-B, CFTR, NKCC1, NCC, KCC, ENaC, and ROMK1, which are mediated by the WNK/SPAK pathway. Additionally, WNKs and SPAK/OSR1 likely control the activation or inhibition of these cotransporters by determining their surface expression level and thereby affecting their trafficking. This information is essential in understanding the entire functionality of PP1 as one of the most important regulation factors of the electrolyte transport. At this point, we are beginning to discover the relationship between PP1 and the WNK/SPAK pathways with respect to regulating epithelial fluid and electrolyte transport. The present study demonstrates the likelihood that the PP1 and NBCe1-B complex control a part of the regulatory mechanism that determines the electrolyte and epithelial fluid secretion.
Go to :
 XML Download
XML Download