

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
The color of mammalian skin and hair is determined by melanin pigment [1]. Melanin pigment is synthesized in melanocytes and translocated to keratinocytes, protecting skin from hazardous environmental stimuli such as UV radiation [23]. Keratinocytes secrete α-melanocyte-stimulating hormone (α-MSH) to protect DNA damage caused by UV radiation [4]. α-MSH binds to melanocortin 1 receptor (MC1R) on melanocytes and stimulates the expression of microphthalmia-associated transcription factor (MITF) through cAMP-dependent pathway, which leads to melanogenesis [5]. However, excessive production of melanin results in hyperpigmentation, such as freckles, melasma, and dark spots [6]. Therefore, modulation of melanogenesis is a critical strategy for managing issues associated with abnormal skin pigmentation [7]. Melanin is synthesized via an enzymatic cascade controlled by tyrosinase, tyrosinase-related protein 1 (TRP1), and TRP2, leading to conversion of tyrosine to melanin pigments [8].
MITF is a central factor in melanogenesis that upregulates expression of tyrosinase, TRP1, and TRP2 [5]. MITF is regulated by transcription factors including MITF itself, cAMP-responsive element binding protein (CREB), paired box gene 3 (PAX3), sex determining region Y-box 10 (SOX10), lymphoid enhancer-binding factor 1/T-cell factor (LEF1/TCF), one cut domain 2 (ONECUT-2), and the mitogen-activated protein kinase (MAPK) pathway [59101112]. Recently, Wnt/β-catenin signal pathway was reported to play a significant role in melanin synthesis [1314]. Wnt ligand binds to cell surface Frizzled receptors, stabilizing β-catenin levels in the cell and ultimately resulting in interaction of β-catenin with LEF1/TCF and activation of the MITF promoter [15].
The various skin whitening agents, such as kojic acid, arbutin, and hydroquinone, have been developed to prevent or treat skin pigmentation. However, some of these agents have revealed harmful side effects, including skin irritation, carcinogenicity, genotoxicity, and oxidative damage to membrane lipids and proteins [1617181920]. For these reasons, some countries have placed restrictions on the use of these agents as cosmetic ingredients. Therefore, there has been an ongoing search for new treatment alternatives.
As stratum corneum, an outermost layer of the epidermis, is the rate-limiting barrier in absorption of an agent, how quickly chemical passes through this outer layer determines epidermis absorption [21]. Lipid-soluble chemicals penetrate the layer and into circulation faster and can be useful as a topical agent to prevent hyperpigmentary disorders such as melasma [222324]. Free fatty acids have been revealed to have regulatory effects on melanogenesis. For example, unsaturated fatty acid such as oleic acid (C18:1), linoleic acid (C18:2), and α-linolenic acid (C18:3) suppress melanin synthesis and tyrosinase activity, whereas saturated fatty acids such as palmitic acid (C16:0) and stearic acid (C18:0) induce melanin synthesis [252627].
Ethyl linoleate (linoleic acid ethyl ester), an unsaturated fatty acid resulting from formal condensation of the carboxyl group of linoleic acid with the hydroxyl group of ethanol, is used in many cosmetics for its antibacterial and anti-inflammatory properties [2829] and is reported to accelerate healing of wounds and clinically proven to be an effective anti-acne agent [30]. It was reported that the ethyl linoleate isolated from Oxalis triangularis inhibited melanogenesis by inhibiting tyrosinase promoter activity [31]. Though there are a few reports stating the anti-melanogenesis activity of ethyl linoleate, the underlying mechanism for tyrosinase modulation by ethyl linoleate is poorly understood. In this study, we examined the depigmentation effects of ethyl linoleate on α-MSH-induced melanogenesis and investigated the underlying molecular mechanisms of anti-melanogenesis. Understanding the molecular mechanisms of ethyl linoleate involved in depigmentation can lead to development of new skin brightening formulations that utilize Akt/GSK3β/β-catenin signal pathway.
METHODS
Chemicals
Dulbecco's modified Eagle's medium (DMEM), fetal bovine serum (FBS), and 100× penicillin/streptomycin solution were obtained from Invitrogen Inc. (Grand Island, NY, USA). Dimethyl sulfoxide (DMSO) and 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) were bought from Amresco Inc. (Solon, OH, USA). Ethyl linoleate, α-melanocyte-stimulating hormone (α-MSH), L-3,4-dihydroxyphenylalanine (L-DOPA), sodium hydroxide, mushroom tyrosinase, arbutin, kojic acid, and resveratrol were obtained from Sigma Chemical Co. (St. Louis, MO, USA). MITF, tyrosinase, and TRP1 antibodies were obtained from Santa Cruz Biotechnology (Santa Cruz, CA, USA) and β-catenin antibody was purchased from BD (Franklin Lakes, NJ, USA). Akt, p-Akt, GSK3β, p-GSK3β, and β-actin antibodies were purchased from Cell signaling Technology (Beverly, MA, USA). Goat anti-mouse and -rabbit IgG secondary antibodies were obtained from Vector Laboratories (Burlingame, CA, USA).
Cell culture
The B16F10 murine melanoma cells were kindly obtained from Dr. Nam Ho Lee, Jeju National University, Korea. The human dermal fibroblast cells were kindly obtained from Dr. Moonjae Cho, Jeju National University, Korea. Both cell lines were cultured by using the DMEM containing 1% penicillin/streptomycin and 10% heat-inactivated FBS in a humidified atmosphere with 5% CO2 incubator at 37℃.
Cell viability
MTT assay was used to detect cell viability. The cells were seeded at a density of 4×103 cells/well for B16F10 and fibroblast cells on 96 well culture plates and cultured overnight. Cells were treated with various concentration of the ethyl linoleate for 48 h. Then, 20 µl of MTT reagent (5 mg/ml) was added to each well at 37℃ for 4 h. After removing the medium, the formazan crystals dissolved with DMSO (150 µl) and the absorbance was measured immediately at 570 nm using a microplate reader (Tecan, Grodig, Austria).
Melanin content
Melanin content was measured with a previously described method with slight modification [32]. After discarding the medium, the cells were harvested and centrifuged. The obtained pellet was dissolved by 1 N sodium hydroxide (NaOH) containing 10% DMSO at 80℃ for 30 min. The relative melanin production was determined by measuring the absorbance at 475 nm using the microplate reader.
In situ intracellular tyrosinase activity
In situ intracellular tyrosinase activity was conducted as previously reported method with minor modification [33]. Cells were fixed with 4% paraformaldehyde for 40 min, washed with PBS, and permeabilized with 0.1% Triton X-100 for 2 min. After washing the cells with PBS, cells were stained with 2 mM L-DOPA for 2 h at 37℃. Staining was imaged and analyzed using a camera attached to a microscope (Olympus, Essex, UK).
Intracellular tyrosinase activity
Intracellular tyrosinase activity was determined using a modified previously method [34]. Cells were collected and lysed in lysis buffer (100 mM Tris–HCl, pH 8, 250 mM NaCl, 0.5% Nonidet P-40, 1× protease inhibitor cocktail), sonicated several times, and then incubated on ice for 30 min. The lysates were centrifuged for 25 min at 14,000 rpm and 4℃, and the protein concentration of supernatants were determined by measuring with a BCA Protein Assay kit (Pierce, Rockford, IL, USA). The diluted lysates were mixed with 2 mM L-DOPA at 37℃, the absorbance was determined at 490 every 10 min for 1 h by a microplate reader.
Mushroom tyrosinase activity
Mushroom tyrosinase activity was used to investigate direct effects of ethyl linoleate on tyrosinase activity by using commercial tyrosinase isolated from mushrooms. The different diluted samples were mixed with 200 U/ml mushroom tyrosinase and 2 mM L-DOPA. After incubation at 37℃ for 10 min, the tyrosinase activity was determined by using the microplate reader at 490 nm absorbance.
Western blot
Western blot was assessed according to the previously described [35]. Equal amounts of protein (10–40 µg) from the cultures were separated by 10–12% SDS-PAGE and transferred onto PVDF membrane (Millipore, Billerica, MA, USA). Membranes were probed with anti-MITF (1:1,000), tyrosinase (1: 7,000), TRP1 (1: 7,000), Akt (1: 2,000), p-Akt (1: 1,000), GSK3β (1: 2,000), p-GSK3β (1: 1,000), β-catenin (1: 2,000), and β-actin (1: 10,000). The membranes were incubated with the secondary antibody at a 1:5,000 dilutions, and protein bands were detected by using the BS ECL plus kit (Biosesang, Gyeonggi-do, Korea). Relative the band intensity was analyzed using ImageJ analysis software (National Institutes of Health, Bethesda, MD, USA).
RESULTS
Effects of ethyl linoleate on the cell viability and melanin content
Cytotoxicity of a drug is critical if the drug is used either as a cosmetic or as a medicine agent [3637]. To examine the cell safety of ethyl linoleate, we treated B16F10 murine melanoma and human dermal fibroblast cells with various concentration of ethyl linoleate. As shown in Fig. 1A and B, ethyl linoleate revealed no significant cytotoxic effect on both cell types at a concentration of 400 µM. Therefore, we used 400 µM ethyl linoleate for further experiments. To evaluate effects of ethyl linoleate on melanogenesis in B16F10 cells, cells were stimulated by propigmenting agents α-MSH in the presence or absence of ethyl linoleate for 48 h. The excessive melanin production with α-MSH treatment was significantly inhibited by ethyl linoleate co-treatment (Fig. 1C). When cells were cultured in medium containing ethyl linoleate, the stimulation of melanin production by α-MSH was reduced to 30.40% by 400 µM ethyl linoleate. According to these results, ethyl linoleate is a noncytotoxic anti-melanogenesis chemical. Therefore, it was studied to evaluate its molecular mechanisms underlying anti-melanogenesis effect by using B16F10 cells.
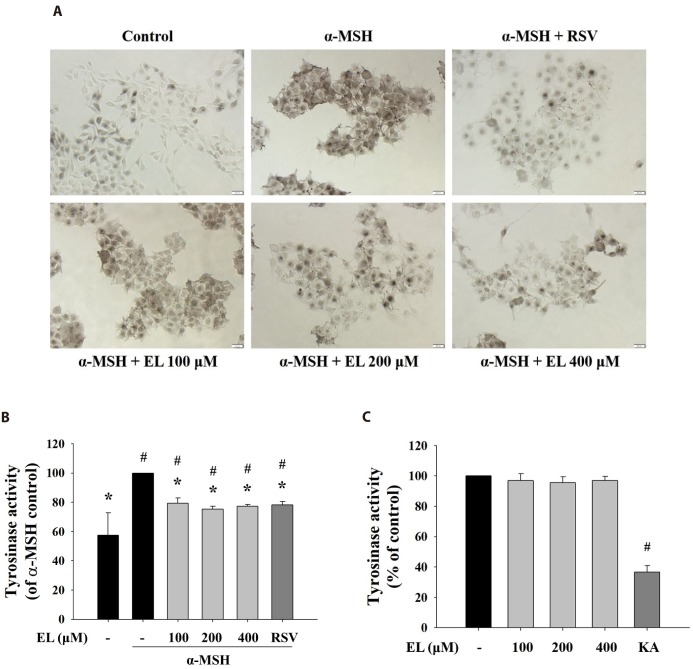
Effects of ethyl linoleate on tyrosinase activity
As tyrosinase is the rate-limiting enzyme critical for melanin biosynthesis, inhibitors of tyrosinase activity have been sought as therapeutic means to treat hyperpigmentary disorders and cosmetic agent [38]. To elucidate ethyl linoleate-mediated inhibition of tyrosinase activity, cells were incubated with L-DOPA, that detected in situ intracellular tyrosinase activity. Treatment with ethyl linoleate for 48 h reduced in situ intracellular tyrosinase activity without affecting cell viability compared to that in the α-MSH control (Fig. 2A). In addition, Fig. 2B revealed that ethyl linoleate treatment induced inhibition of intracellular tyrosinase activity compared to that in the α-MSH control. Whereas, under cell-free conditions, ethyl linoleate revealed no notable inhibitory effect on the activity of mushroom tyrosinase activity (Fig. 2C), suggesting that the effect on B16F10 cells was not mediated by direct interaction of ethyl linoleate with the tyrosinase enzyme. Data indicated that ethyl linoleate probably inhibited the function of tyrosinase by decreasing tyrosinase expression.
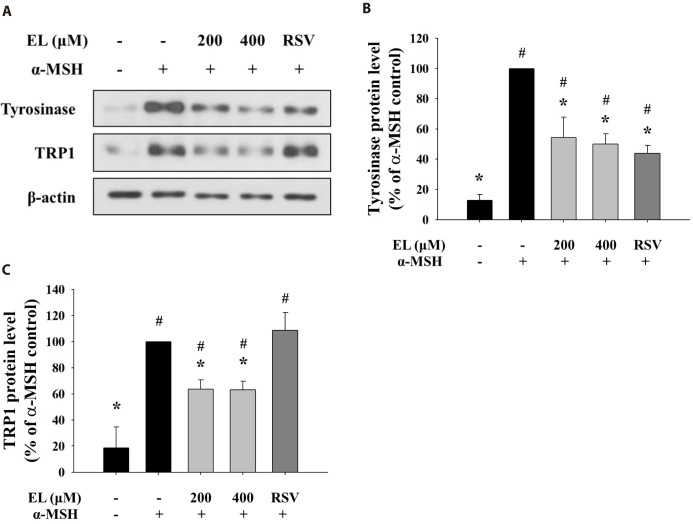
Effects of ethyl linoleate on melanogenic enzyme protein expression
As ethyl linoleate treatment induced inhibition of intracellular tyrosinase activity, not mushroom tyrosinase activity, we determined levels of melanogenic enzyme proteins, such as tyrosinase and tyrosinase-related protein 1 (TRP1). Fig. 3A shows that protein expressions of tyrosinase and TRP1 were repressed with increasing concentration of ethyl linoleate. Quantification of the detected signals revealed that protein levels of tyrosinase and TRP1 following treatment with 400 µM ethyl linoleate were reduced to 50.02% and 63.17% of the α-MSH control, respectively (Figs. 3B and C). While resveratrol, a tyrosinase inhibitor, only reduced expression of tyrosinase, not TRP1, suggesting that ethyl linoleate treatment reduced synthesis of melanin more than resveratrol treatment due to the reduction in the expression of both tyrosinase and TRP1.
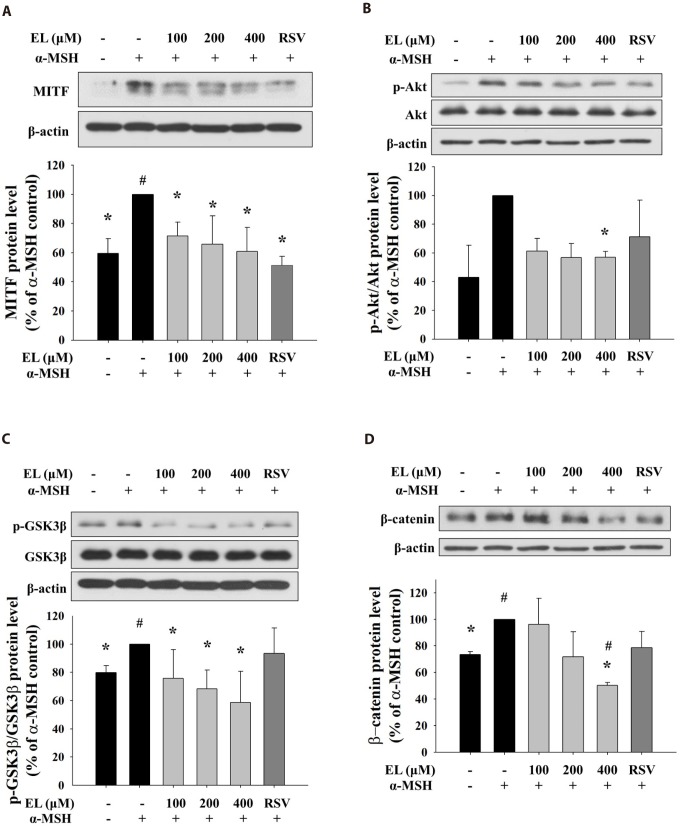
Ethyl linoleate inhibits MITF expression by regulating Akt and GSK3β/β-catenin signal pathway
To evaluate the mechanism of the inhibitory action of ethyl linoleate on melanogenesis, we treated cells with various concentration of ethyl linoleate with α-MSH and examined the involvement of Akt/GSK3β/β-catenin signaling pathway. The microphthalmia-associated transcription factor (MITF) is widely regarded as the key transcriptional regulator of melanogenic enzyme proteins. As shown in Fig. 4A, the expression of MITF was reduced in a dose-dependent manner by 71.49%, 65.73%, and 60.80% in the presence of 100, 200, and 400 µM ethyl linoleate, respectively, compared to α-MSH alone treatment. Ethyl linoleate decreased the ratio of phosphorylated-form/total-form of Akt and GSK3β by 56.96% and 58.54% in the presence of 400 µM ethyl linoleate, respectively, compared to α-MSH alone (Fig. 4B, C). Furthermore, ethyl linoleate inhibited level of expression of β-catenin in a concentration-dependent manner by 96.23%, 71.67%, and 50.31% in the presence of 100, 200, and 400 µM ethyl linoleate, respectively, compared to α-MSH alone treatment (Fig. 4D). According to these results, hyperpigmentation inhibition by ethyl linoleate is associated with inhibition of phosphorylation of Akt and GSK3β that led to suppression of MITF expression through degradation of β-catenin, reduced tyrosinase and TRP1 expression, and inhibited melanin production.
To further confirm involvement of Akt/GSK3β/β-catenin signaling pathway in ethyl linoleate-induced inhibition of melanogenesis, we conducted a time course experiment to establish the kinetic of melanogenesis-related proteins expression following 400 µM ethyl linoleate treatment in α-MSH stimulation. As shown in supplementary Fig. 1, α-MSH treatment markedly increased levels of tyrosinase and TRP1 in a time-dependent manner. MITF expression was increased after α-MSH treatment, which peaked at 4 h, followed by a continuous decline to 24 h, and a recovery to basal levels after 48 h of treatment. The ethyl linoleate co-treatment with α-MSH began to decrease the protein level of MITF at 4 h, which continued decline to 12 h. In addition, we observed that the phosphorylated Akt by ethyl linoleate rapidly declined more than those by α-MSH only. The ethyl linoleate cotreatment with α-MSH caused decrease in the phosphorylation of GSK3β levels; however, there was no significant reduction in the phosphorylation of GSK3β following the α-MSH alone treatment. It was also apparent that ethyl linoleate co-treatment with α-MSH resulted in the loss of β-catenin expression from 4 h to 48 h.
DISCUSSION
Many investigations have focused on the specific mechanisms involved in melanogenesis to develop new therapeutic agents for skin pigmentation abnormalities. Pigmentation may be regulated by different steps: expression of melanogenic enzyme proteins, regulation of melanogenic enzyme activity during or before melanin synthesis, melanosome transfer to recipient keratinocytes, and melanosome degradation and turnover. Among these steps, the expression of melanogenic enzyme proteins is the most important step for regulation of pigmentation [39]. Tyrosinase, tyrosinase-related protein 1 (TRP1), and TRP2 are key enzymes for melanin biosynthesis, and these enzymes are influenced by microphthalmia-associated transcription factor (MITF) [5].
Previous studies have been reported that unsaturated fatty acids, such as oleic acid, linoleic acid, and α-linolenic acid, suppress melanin biosynthesis through the inhibition of tyrosinase activities. It has revealed that linoleic acid enhances the ubiquitination of mature tyrosinase and the ubiquitinated tyrosinase could be integrated in the ER-associated degradation after rapid processing of tyrosinase from the ER to the Golgi [2627]. However, the underlying mechanism for tyrosinase modulation by ethyl linoleate is poorly understood thus far. In the present study, we observed that ethyl linoleate decreased the expression of tyrosinase and TRP1 through the reduction of MITF expression as shown in Fig. 4. Furthermore, we found that hyperpigmentation inhibition by ethyl linoleate is associated with inhibition of phosphorylation of Akt and GSK3β that led to suppression of MITF expression through degradation of β-catenin in a concentration dependent manner.
It is known that MITF is regulated by the balance between a variety of signal transduction pathway, including the cyclic adenosine monophosphate/protein kinase A (cAMP/PKA), extracellular signal-regulated kinase (ERK), p38, c-Jun N-terminal kinase (JNK), and Akt pathway [1240414243]. While we could not detect changes in phosphorylation of CREB, ERK, JNK, and p38 (data not shown), phosphorylation of Akt was decreased compared to α-MSH control (Fig. 4B). It has been reported that activated Akt can phosphorylate glycogen synthase kinase 3β (GSK3β) [44]. When GSK3β is phosphorylated, GSK3β-dependent phosphorylation of β-catenin is blocked and β-catenin is translocated into the nucleus, that recruits the complex of β-catenin and LEF1/TCF to binding sites of the MITF promoter [1445]. Our data showed that ethyl linoleate treatment decreased the expression of p-Akt, p-GSK3β, and β-catenin (Figs. 4B-D). Therefore, the antimelanogenesis effect by ethyl linoleate contributed to suppression of MITF expression through Akt/GSK3β/β-catenin signaling pathway.
In conclusion, ethyl linoleate decreased melanin production and tyrosinase activity through the reduction of tyrosinase and TRP1 expression. Our data revealed that ethyl linoleate treatment decreased expression of MITF through Akt/GSK3β/β-catenin pathway. This study provides a molecular basis for understanding inhibitory effects of ethyl linoleate on melanogenesis. Ethyl linoleate may be used as a non-cytotoxic and skin-whitening agent as a cosmetic and medicine.

 XML Download
XML Download