

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Epilepsy, a chronic brain disorder, affects approximately 2% of the population worldwide [1]. Currently available antiepileptic drugs are all synthetic compounds, which are associated with major therapeutic problems such as serious side effects and drug resistance [234]. Therefore, the discovery and development of new antiepileptic drugs with higher efficacy and fewer side effects are still desirable. Plants are a rich source of novel compounds for developing new drugs. Numerous herbal constituents have shown promising results in treating experimental and clinical epilepsy [56], and many of these natural compounds appear to be safe. Echinacoside is an active compound in the herb Herba Cistanche [7], which is used as a traditional Chinese medicine for the treatment of epilepsy [89]. Echinacoside enters the brain and produces numerous beneficial effects, including neuroprotection and cognition improvement [101112]. In our previous study, we demonstrated that echinacoside inhibits the release of the excitatory neurotransmitter glutamate from cortical nerve endings [13]. Because glutamate plays a key role in epilepsy and antiepileptic drug therapy [14], we hypothesized that echinacoside has anticonvulsant properties. To address this supposition, the present study used an electrophysiological approach in rat hippocampal slices to evaluate the effects of echinacoside on epileptiform activity induced by 4-aminopyridine (4-AP), which is commonly used for epileptogenesis in vitro [1516]. In addition, because it has been suggested that the blockade of sustained repetitive firing in neurons is one of the basic mechanisms of antiepileptic drugs [1718], we examined the effect of echinacoside on sustained repetitive firing in hippocampal pyramidal neurons.
Go to : 
METHODS
Animals
We purchased 14- to 23-day-old male Sprague–Dawley rats from BioLASCO Taiwan Co., Ltd. (Taipei, Taiwan). The rats were housed at a constant temperature (22±1℃) and relative humidity (50%), under a regular 12-h light–dark cycle (lights off at 7 pm). Food and water were freely available. The experimental procedures were approved by the Fu Jen Institutional Animal Care and Utilization Committee (IACUC No. A10259), in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals. All efforts were made to minimize animal suffering and to use a minimum number of animals necessary to produce reliable results.
Slice preparation
Hippocampal slices were prepared from 14- to 23-day-old male rats for whole-cell patch clamp recordings, through procedures described previously [19]. Briefly, after decapitation, the hippocampus was positioned on the stage of a vibratome slicer (VT1000S, Leica, Germany) and cut to obtain 300-µm-thick transverse brain slices. The slices were maintained in artificial cerebrospinal fluid continuously oxygenated with 95% O2-5% CO2 at room temperature for at least 1 h before use. Artificial cerebrospinal fluid comprised 117 mM NaCl, 4.7 mM KCl, 2.5 mM CaCl2, 1.2 mM MgCl2, 25 mM NaHCO3, 1.2 mM NaH2PO4, and 11 mM glucose (pH 7.4, 300 mOsm).
Electrophysiological recordings
Slices were moved to a recording chamber mounted on a BX51W1 upright microscope (Olympus; Tokyo, Japan) equipped with infrared differential interference contrast. The chamber was continuously perfused with oxygenated artificial cerebrospinal fluid. Neurons were visualized using an Olympus Optical 40× water immersion objective. Whole-cell voltage clamp or current clamp recordings in hippocampal CA3 neurons were performed using pipettes of 2–5 MΩ resistance after files with an internal solution containing 0.3 mM Na3GTP, 135 mM K-gluconate, 20 mM KCl, 0.1 mM EGTA, 2 mM MgCl2, 4 mM Na2ATP, and 10 mM HEPES (pH 7.3, 280 mOsm). Pipettes were fabricated from borosilicate glass (outer diameter, 1.5 mm; internal diameter, 0.86 mm) by using a micropipette puller (P87, Sutter Instruments, USA). Positive pressure was applied to the recording pipette as it was lowered into the medium and approached the cell membrane. A constant negative pressure was applied to form the seal (>1 GΩ) when the recording pipette was attached to the membrane, and the cell membrane was then drawn up quickly to rupture it and access the whole-cell configuration. Neurons were voltage clamped at −70 mV by using an Axopatch 200B amplifier (Axon Instruments, Foster City, CA, USA). Access resistance (8–30 MΩ) was regularly monitored during recordings, and cells were rejected if the resistance changed by >20% during the experiments. Membrane currents were filtered at 2 kHz, digitized, and stored on a computer using pCLAMP (Axon Instruments, Foster City, CA, USA). Data were analyzed offline using commercially available software (Synaptosoft Inc., Fort Lee, NJ, USA).
Data analysis
All data are expressed as means±SEM and were compared using Student's t-test. p<0.05 was considered significant.
Go to :
RESULTS
Echinacoside suppresses epileptiform activity induced by 4-AP
Spontaneous epileptiform discharges were observed in the hippocampal CA3 pyramidal neurons after 10 min of perfusion with 1 mM 4-AP, as described previously [1720]. The spontaneous bursts that occurred at a frequency of 1.05±0.23 Hz had durations of 22.0±2.43 s and was observed 30±4 times (Fig. 1A). The addition of 20 µM echinacoside shortened the duration of burst discharges. As shown in Fig. 1A, 10 min after introduction of echinacoside (20 µM) into the perfusion chamber, the burst duration was decreased by an average of 82.6%±5.2% (3.6±0.75 s, n=5; p<0.001). The effect of echinacoside was dose dependent, with an IC50 of approximately 20 µM (Fig. 1B). Echinacoside had no effect in the control conditions (Fig. 1A).
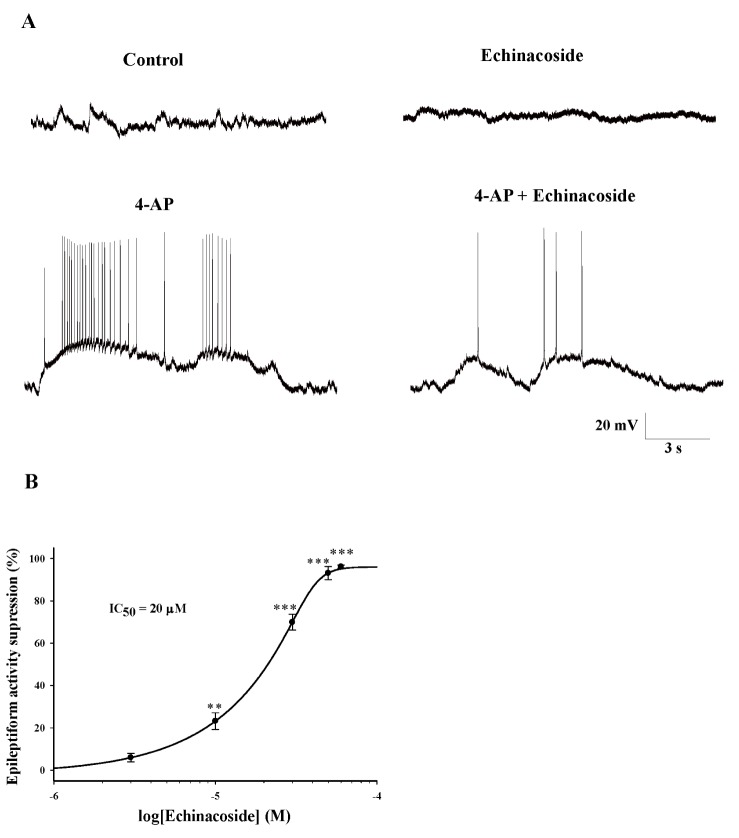 | Fig. 1Effect of echinacoside on the epileptiform activity induced by 4-AP in CA3 pyramidal neurons in rat hippocampal slices.(A) Representative current clamp recordings in control, echinacoside (20 µM), 4-AP (1 mM), and 4-AP (1 mM)+echinacoside (20 µM). 4-AP was applied for 10 min to induce epileptiform activity. Echinacoside was added 10 min after the application of 4-AP. (B) Normalized dose-response relationship of the effect of echinacoside on epileptiform activity induced by 4-AP (n=3-6). **p<0.01, ***p<0.001 vs. control group.
|
Echinacoside decreases the frequency but not the amplitude of spontaneous excitatory postsynaptic currents induced by 4-AP
Studies have suggested that epileptiform activity induced by 4-AP in the hippocampal CA3 pyramidal neurons is associated with an increase of spontaneous excitatory synaptic activity [1621]. Therefore, we examined the effects of echinacoside on spontaneous excitatory postsynaptic currents (sEPSCs) under 4-AP conditions. sEPSCs were recorded at a holding potential of −70 mV and in the presence of the GABAA receptor antagonist bicuculline (20 µM). Very few sEPSCs were observed in the hippocampal CA3 pyramidal neurons in control conditions (Fig. 2A). Under control conditions, the bath application of 20 µM echinacoside for 10 min reduced the sEPSC frequency (p<0.001), without changing the amplitude (p=0.16) (n=5). Perfusion with 1 mM 4-AP for 10 min induced a significant increase in the frequency and amplitude of sEPSCs (Fig. 2A; p<0.001). After 10 min perfusion with 20 µM echinacoside, the increase in sEPSC frequency induced by 4-AP was reduced (Figs. 2A and B; n=5; p<0.05), whereas the sEPSC amplitude was not significantly affected (Figs. 2A and C; n=5; p=0.98).
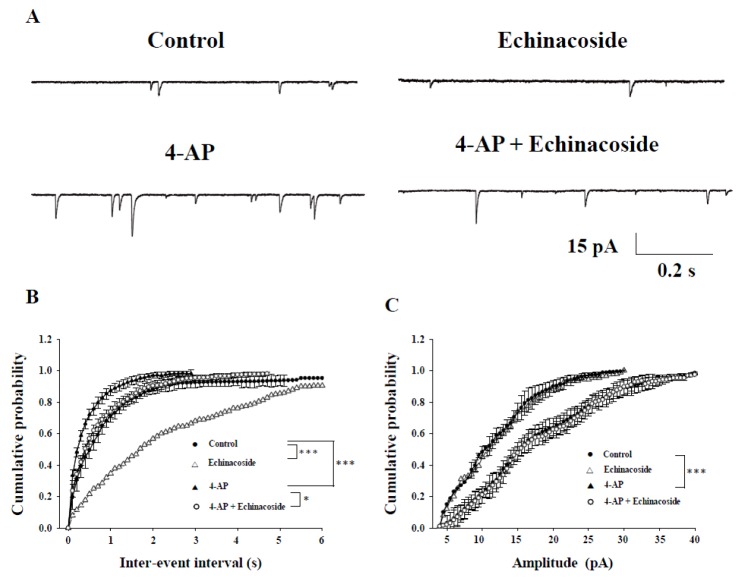 | Fig. 2Effect of echinacoside on spontaneous excitatory postsynaptic potentials (sEPSCs) induced by 4-AP in hippocampal CA3 pyramidal neurons.(A) Representative voltage clamp recordings in control, echinacoside (20 µM), 4-AP (1 mM), and 4-AP (1 mM)+echinacoside (20 µM). Echinacoside was added 10 min after the application of 4-AP. (B, C) Cumulative probability plots of sEPSCs frequency and amplitude, respectively. Each result represents mean values from 5 neurons. ***p<0.001 vs. control group. *p<0.05 vs. the 4-AP-treated group.
|
In addition, miniature excitatory postsynaptic currents (mEPSCs) were recorded at a holding potential of −70 mV and in the presence of the Na+ channel blocker tetrodotoxin (TTX, 1 µM) and the GABAA receptor antagonist bicuculline (20 µM). As shown in Fig. 3A, application of 20 µM echinacoside for 10 min reduced the frequency of mEPSCs (Figs. 3A and B; n=5; p<0.001) without affecting their amplitude (Figs. 3A and C; n=5; p=0.84).
 | Fig. 3Effect of echinacoside on miniature excitatory postsynaptic potentials (mEPSCs) in hippocampal CA3 pyramidal neurons.(A) Representative voltage clamp recordings from neurons before (control) and after echinacoside (20 µM) application for 10 min. mEPSCs recorded in the presence of TTX. (B, C) Cumulative probability plots of mEPSCs frequency and amplitude, respectively. Each result represents mean values from 5 neurons. ***p<0.001 vs. control group.
|
Echinacoside does not affect postsynaptic glutamate receptor sensitivity
We examined the effect of echinacoside on the postsynaptic responses to exogenously applied 100 µM glutamate. The experiments were performed in the presence of TTX (0.5 µM). As illustrated in Fig. 4A, after 10 min perfusion with 20 µM echinacoside, the amplitude of the evoked glutamate current (IGlu) was not significant different from that in the control condition (control, 1320±97 pA; echinacoside, 1240±117 pA; n=5; p=0.61; Fig. 4B).
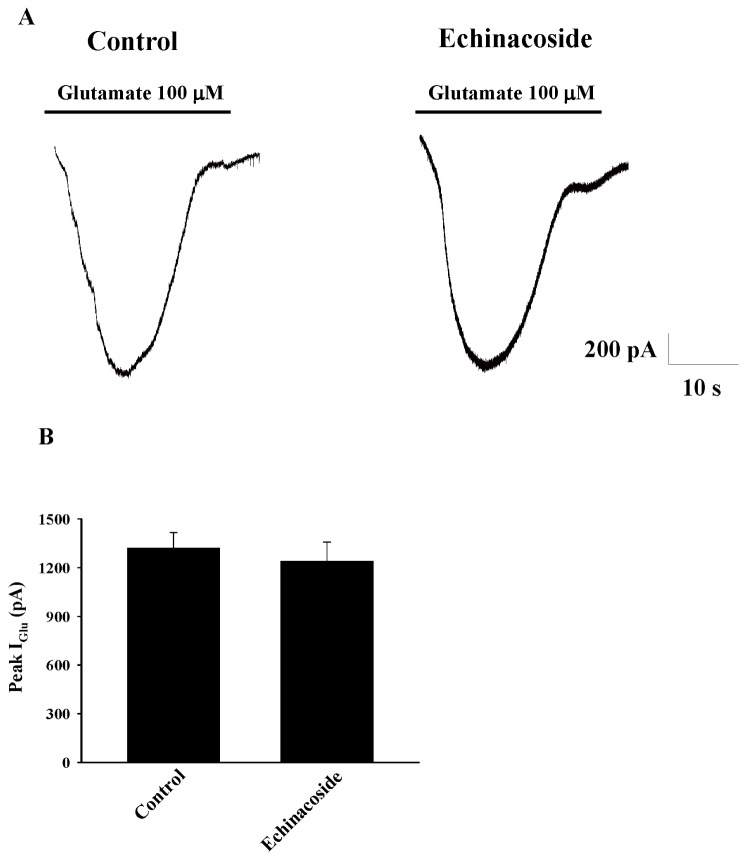 | Fig. 4Effect of echinacoside on glutamate-evoked postsynaptic currents (IGlu) in hippocampal CA3 pyramidal neurons.Superfusion of 100 µM glutamate for 10 s evoked an inward current. (A) Typical IGlu traces recorded from a CA3 pyramidal neuron before (control) and after echinacoside (20 µM) application for 10 min. (B) Summary data (n=5) showing mean values of peak IGlu in the absence (control) and presence of 20 µM echinacoside.
|
Echinacoside reduces the number of sustained repetitive action potentials
In addition, because the blockade of sustained repetitive firing in neurons has been suggested to be a basic mechanisms of antiepileptic drugs [1718], we examined the effect of echinacoside on sustained repetitive firing in hippocampal CA3 pyramidal neurons. As shown in Fig. 5A, sustained repetitive firing was induced by a 500-ms, 0.3-Hz, 1-nA depolarizing pulse. Perfusion with 20 µM echinacoside for 10 min decreased the frequency of the action potentials induced by pulse depolarization (control, 18.57±0.8 Hz; echinacoside, 8.71±05 Hz; n=5; p=0.000; Figs. 5A and B).
 | Fig. 5Effect of echinacoside on sustained repetitive firing in hippocampal CA3 pyramidal neurons.(A) Representative current clamp recordings of action potential induced by depolarizing current pulses (500 ms; 1 nA) from neurons before (control) and after echinacoside (20 µM) application for 10 min. (B) Summary data (n=7) showing mean values of action potential frequency in the absence (control) and presence of 20 µM echinacoside. ***p<0.001 vs. control group.
|
Go to :
DISCUSSION
Excessive glutamate release has been proposed to be involved in the pathogenesis of epilepsy [22], and decreasing the brain glutamate levels is considered to be a treatment strategy for this disorder. We previously showed that echinacoside, an active compound in herb Herba Cistanche, inhibits glutamate release in cortical nerve terminals [13]. Accordingly, in this study, the potential antiepileptic activity of echinacoside was investigated using an in vitro model in which epileptiform activity in rat hippocampal slices was induced by 4-AP. In accordance with the results of previous studies [1720], the perfusion of 1 mM 4-AP in the hippocampal CA3 pyramidal neurons resulted in epileptiform activity. This 4-AP-induced epileptiform activity was suppressed by echinacoside at concentrations between 5 and 30 µM, suggesting that echinacoside exerts an antiepileptic effect.
Previous studies have suggested that 4-AP-induced epileptiform activity is associated with an increase in spontaneous excitatory synaptic activity in hippocampal slices [1621]. In the present study, we also observed that 4-AP caused a significant increase in the frequency and amplitude of sEPSCs in the hippocampal CA3 pyramidal neurons. However, the increased frequency of sEPSCs induced by 4-AP was reduced by echinacoside, with no change in amplitude. Similar effects were seen on mEPSCs, recorded in the presence of TTX. The change observed in the frequency of sEPSCs and mEPSCs is a result of presynaptic action, whereas the change in their amplitude typically indicates postsynaptic modulation [232425]. The lack of effect on the amplitude of sEPSCs and mEPSCs observed in our study indicates that echinacoside does not block postsynaptic glutamate receptors, but the decrease in the frequency preferentially indicates a presynaptic inhibition of glutamate release. This suggestion was supported by our observation that postsynaptic responses to glutamate was not significantly altered by echinacoside. Furthermore, echinacoside has been shown to inhibit the release of glutamate from cortical nerve terminals [13]. Thus, we suggested that echinacoside suppresses 4-AP-induced epileptiform activity by decreasing presynaptic glutamate release in the hippocampal CA3 pyramidal neurons.
In addition, we observed that echinacoside profoundly blocked the sustained repetitive firing of action potentials in the hippocampal CA3 pyramidal neurons. This action on sustained repetitive firing was similar to the effects of the currently available antiepileptic drugs phenytoin and topiramate [1826], indicating that echinacoside blocks abnormal firing synchronization among pyramidal neurons. Although the mechanism underlying the antiepileptic effect of echinacoside requires further investigation, this finding suggests that echinacoside may prevent the spread of seizure discharges during epileptic seizures.
In conclusion, the present results demonstrate that echinacoside effectively reduces spontaneous glutamate release and blocks sustained repetitive firing of action potentials in hippocampal CA3 pyramidal neurons. These reciprocal effects are highly desirable actions required in an effective antiepileptic drug.
Go to :
 XML Download
XML Download