1. García AC, de Souza LG, Pereira MG, Castro RN, García-Mina JM, Zonta E, et al. Structure-property-function relationship in humic substances to explain the biological activity in plants. Sci Rep. 2016; 6:20798. PMID:
26862010.

2. Sehrawat A, Roy R, Pore SK, Hahm ER, Samanta SK, Singh KB, Kim SH, Singh K, Singh SV. Mitochondrial dysfunction in cancer chemoprevention by phytochemicals from dietary and medicinal plants. Semin Cancer Biol. 2017; 47:147–153. PMID:
27867044.
3. Diarra M, El Ouahabi H, Bouxid H, Boujraf S, Khabbal Y, Ajdi F. Medicinal plants in type 2 diabetes: therapeutic and economical aspects. Int J Prev Med. 2016; 7:56. PMID:
27076894.
4. de Freitas Junior LM, de Almeida EB Jr. Medicinal plants for the treatment of obesity: ethnopharmacological approach and chemical and biological studies. Am J Transl Res. 2017; 9:2050–2064. PMID:
28559960.
5. Croft KD, Yamashita Y, O'Donoghue H, Shirasaya D, Ward NC, Ashida H. Screening plant derived dietary phenolic compounds for bioactivity related to cardiovascular disease. Fitoterapia. 2017; DOI:
10.1016/j.fitote.2017.12.002. [Epub ahead of print].
6. Jiang T, Wang XQ, Ding C, Du XL. Genistein attenuates isofluraneinduced neurotoxicity and improves impaired spatial learning and memory by regulating cAMP/CREB and BDNF-TrkB-PI3K/Akt signaling. Korean J Physiol Pharmacol. 2017; 21:579–589. PMID:
29200900.
7. Leake I. Liver: plant sterols have a role in liver injury associated with parenteral nutrition. Nat Rev Gastroenterol Hepatol. 2013; 10:693.
8. Katashima CK, Silva VR, Gomes TL, Pichard C, Pimentel GD. Ursolic acid and mechanisms of actions on adipose and muscle tissue: a systematic review. Obes Rev. 2017; 18:700–711. PMID:
28335087.
9. Kunkel SD, Suneja M, Ebert SM, Bongers KS, Fox DK, Malmberg SE, Alipour F, Shields RK, Adams CM. mRNA expression signatures of human skeletal muscle atrophy identify a natural compound that increases muscle mass. Cell Metab. 2011; 13:627–638. PMID:
21641545.
10. Woźniak Ł, Skąpska S, Marszałek K. Ursolic acid—a pentacyclic triterpenoid with a wide spectrum of pharmacological activities. Molecules. 2015; 20:20614–20641. PMID:
26610440.
11. Jäger S, Trojan H, Kopp T, Laszczyk MN, Scheffler A. Pentacyclic triterpene distribution in various plants - rich sources for a new group of multi-potent plant extracts. Molecules. 2009; 14:2016–2031. PMID:
19513002.
12. Hussain H, Green IR, Ali I, Khan IA, Ali Z, Al-Sadi AM, Ahmed I. Ursolic acid derivatives for pharmaceutical use: a patent review (2012-2016). Expert Opin Ther Pat. 2017; 27:1061–1072. PMID:
28637397.
13. Mancha-Ramirez AM, Slaga TJ. Ursolic acid and chronic disease: an overview of UA's effects on prevention and treatment of obesity and cancer. Adv Exp Med Biol. 2016; 928:75–96. PMID:
27671813.
14. Kashyap D, Sharma A, Tuli HS, Punia S, Sharma AK. Ursolic acid and oleanolic acid: pentacyclic terpenoids with promising antiinflammatory activities. Recent Pat Inflamm Allergy Drug Discov. 2016; 10:21–33. PMID:
27531153.
15. Liobikas J, Majiene D, Trumbeckaite S, Kursvietiene L, Masteikova R, Kopustinskiene DM, Savickas A, Bernatoniene J. Uncoupling and antioxidant effects of ursolic acid in isolated rat heart mitochondria. J Nat Prod. 2011; 74:1640–1644. PMID:
21648406.
16. Shishodia S, Majumdar S, Banerjee S, Aggarwal BB. Ursolic acid inhibits nuclear factor-kappaB activation induced by carcinogenic agents through suppression of IkappaBalpha kinase and p65 phosphorylation: correlation with down-regulation of cyclooxygenase 2, matrix metalloproteinase 9, and cyclin D1. Cancer Res. 2003; 63:4375–4383. PMID:
12907607.
17. Jayaprakasam B, Olson LK, Schutzki RE, Tai MH, Nair MG. Amelioration of obesity and glucose intolerance in high-fat-fed C57BL/6 mice by anthocyanins and ursolic acid in Cornelian cherry (Cornus mas). J Agric Food Chem. 2006; 54:243–248. PMID:
16390206.
18. Yu SG, Zhang CJ, Xu XE, Sun JH, Zhang L, Yu PF. Ursolic acid derivative ameliorates streptozotocin-induced diabestic bone deleterious effects in mice. Int J Clin Exp Pathol. 2015; 8:3681–3690. PMID:
26097549.
19. Senthil S, Chandramohan G, Pugalendi KV. Isomers (oleanolic and ursolic acids) differ in their protective effect against isoproterenolinduced myocardial ischemia in rats. Int J Cardiol. 2007; 119:131–133. PMID:
17052790.
20. Wang Y, He Z, Deng S. Ursolic acid reduces the metalloprotease/ anti-metalloprotease imbalance in cerebral ischemia and reperfusion injury. Drug Des Devel Ther. 2016; 10:1663–1674.
21. Sundaresan A, Radhiga T, Pugalendi KV. Effect of ursolic acid and Rosiglitazone combination on hepatic lipid accumulation in high fat diet-fed C57BL/6J mice. Eur J Pharmacol. 2014; 741:297–303. PMID:
25149666.
22. Bakhtiari N, Hosseinkhani S, Soleimani M, Hemmati R, Noori-Zadeh A, Javan M, Tashakor A. Short-term ursolic acid promotes skeletal muscle rejuvenation through enhancing of SIRT1 expression and satellite cells proliferation. Biomed Pharmacother. 2016; 78:185–196. PMID:
26898441.
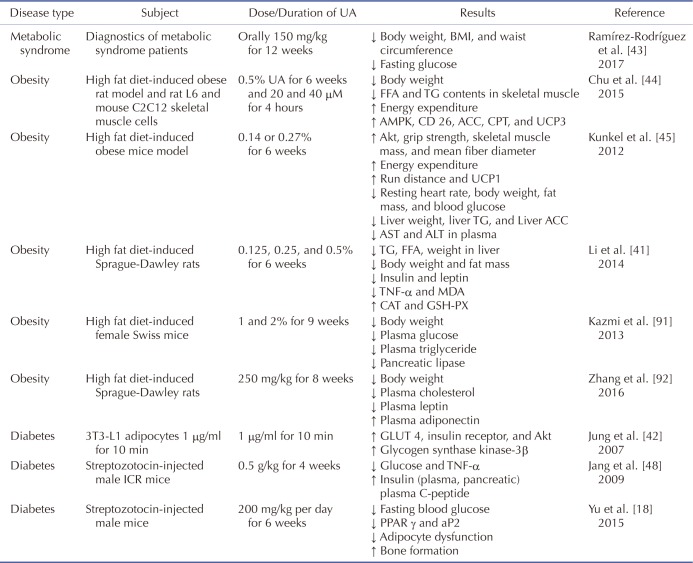
23. Kashyap D, Tuli HS, Sharma AK. Ursolic acid (UA): a metabolite with promising therapeutic potential. Life Sci. 2016; 146:201–213. PMID:
26775565.
24. Huang MT, Ho CT, Wang ZY, Ferraro T, Lou YR, Stauber K, Ma W, Georgiadis C, Laskin JD, Conney AH. Inhibition of skin tumorigenesis by rosemary and its constituents carnosol and ursolic acid. Cancer Res. 1994; 54:701–708. PMID:
8306331.
25. Prasad S, Yadav VR, Sung B, Reuter S, Kannappan R, Deorukhkar A, Diagaradjane P, Wei C, Baladandayuthapani V, Krishnan S, Guha S, Aggarwal BB. Ursolic acid inhibits growth and metastasis of human colorectal cancer in an orthotopic nude mouse model by targeting multiple cell signaling pathways: chemosensitization with capecitabine. Clin Cancer Res. 2012; 18:4942–4953. PMID:
22832932.
26. Lewinska A, Adamczyk-Grochala J, Kwasniewicz E, Deregowska A, Wnuk M. Ursolic acid-mediated changes in glycolytic pathway promote cytotoxic autophagy and apoptosis in phenotypically different breast cancer cells. Apoptosis. 2017; 22:800–815. PMID:
28213701.
27. Li W, Zhang H, Nie M, Tian Y, Chen X, Chen C, Chen H, Liu R. Ursolic acid derivative FZU-03,010 inhibits STAT3 and induces cell cycle arrest and apoptosis in renal and breast cancer cells. Acta Biochim Biophys Sin (Shanghai). 2017; 49:367–373. PMID:
28338932.
28. Luo J, Hu YL, Wang H. Ursolic acid inhibits breast cancer growth by inhibiting proliferation, inducing autophagy and apoptosis, and suppressing inflammatory responses via the PI3K/AKT and NF-κB signaling pathways in vitro. Exp Ther Med. 2017; 14:3623–3631. PMID:
29042957.
29. Cao C, Wang W, Lu L, Wang L, Chen X, Guo R, Li S, Jiang J. Inactivation of Beclin-1-dependent autophagy promotes ursolic acidinduced apoptosis in hypertrophic scar fibroblasts. Exp Dermatol. 2018; 27:58–63. PMID:
28767174.
30. Elstrom RL, Bauer DE, Buzzai M, Karnauskas R, Harris MH, Plas DR, Zhuang H, Cinalli RM, Alavi A, Rudin CM, Thompson CB. Akt stimulates aerobic glycolysis in cancer cells. Cancer Res. 2004; 64:3892–3899. PMID:
15172999.
31. Robey RB, Hay N. Is Akt the “Warburg kinase”?-Akt-energy metabolism interactions and oncogenesis. Semin Cancer Biol. 2009; 19:25–31. PMID:
19130886.
32. Zheng QY, Jin FS, Yao C, Zhang T, Zhang GH, Ai X. Ursolic acidinduced AMP-activated protein kinase (AMPK) activation contributes to growth inhibition and apoptosis in human bladder cancer T24 cells. Biochem Biophys Res Commun. 2012; 419:741–747. PMID:
22387548.
33. Shen S, Zhang Y, Zhang R, Tu X, Gong X. Ursolic acid induces autophagy in U87MG cells via ROS-dependent endoplasmic reticulum stress. Chem Biol Interact. 2014; 218:28–41. PMID:
24802810.
34. Xavier CP, Lima CF, Pedro DF, Wilson JM, Kristiansen K, Pereira-Wilson C. Ursolic acid induces cell death and modulates autophagy through JNK pathway in apoptosis-resistant colorectal cancer cells. J Nutr Biochem. 2013; 24:706–712. PMID:
22841540.
35. Leng S, Hao Y, Du D, Xie S, Hong L, Gu H, Zhu X, Zhang J, Fan D, Kung HF. Ursolic acid promotes cancer cell death by inducing Atg5-dependent autophagy. Int J Cancer. 2013; 133:2781–2790. PMID:
23737395.
36. Li J, Liang X, Yang X. Ursolic acid inhibits growth and induces apoptosis in gemcitabine-resistant human pancreatic cancer via the JNK and PI3K/Akt/NF-κB pathways. Oncol Rep. 2012; 28:501–510. PMID:
22641480.
37. Song YH, Jeong SJ, Kwon HY, Kim B, Kim SH, Yoo DY. Ursolic acid from Oldenlandia diffusa induces apoptosis via activation of caspases and phosphorylation of glycogen synthase kinase 3 beta in SKOV-3 ovarian cancer cells. Biol Pharm Bull. 2012; 35:1022–1028. PMID:
22791147.
38. Yan SL, Huang CY, Wu ST, Yin MC. Oleanolic acid and ursolic acid induce apoptosis in four human liver cancer cell lines. Toxicol In Vitro. 2010; 24:842–848. PMID:
20005942.
39. Chen JC, Chung JG, Chen LD. Gypenoside induces apoptosis in human Hep3B and HA22T tumour cells. Cytobios. 1999; 100:37–48. PMID:
10643643.
40. Nagamine T, Hayakawa K, Kusakabe T, Takada H, Nakazato K, Hisanaga E, Iha M. Inhibitory effect of fucoidan on Huh7 hepatoma cells through downregulation of CXCL12. Nutr Cancer. 2009; 61:340–347. PMID:
19373607.
41. Li S, Liao X, Meng F, Wang Y, Sun Z, Guo F, Li X, Meng M, L Y, Sun C. Therapeutic role of ursolic acid on ameliorating hepatic steatosis and improving metabolic disorders in high-fat diet-induced nonalcoholic fatty liver disease rats. PLoS One. 2014; 9:e86724. PMID:
24489777.
42. Jung SH, Ha YJ, Shim EK, Choi SY, Jin JL, Yun-Choi HS, Lee JR. Insulin-mimetic and insulin-sensitizing activities of a pentacyclic triterpenoid insulin receptor activator. Biochem J. 2007; 403:243–250. PMID:
17201692.
43. Ramírez-Rodríguez AM, González-Ortiz M, Martínez-Abundis E, Acuña Ortega N. Effect of ursolic acid on metabolic syndrome, insulin sensitivity, and inflammation. J Med Food. 2017; 20:882–886. PMID:
28598231.
44. Chu X, He X, Shi Z, Li C, Guo F, Li S, Li Y, Na L, Sun C. Ursolic acid increases energy expenditure through enhancing free fatty acid uptake and β-oxidation via an UCP3/AMPK-dependent pathway in skeletal muscle. Mol Nutr Food Res. 2015; 59:1491–1503. PMID:
25944715.
45. Kunkel SD, Elmore CJ, Bongers KS, Ebert SM, Fox DK, Dyle MC, Bullard SA, Adams CM. Ursolic acid increases skeletal muscle and brown fat and decreases diet-induced obesity, glucose intolerance and fatty liver disease. PLoS One. 2012; 7:e39332. PMID:
22745735.
46. Jia Y, Bhuiyan MJ, Jun HJ, Lee JH, Hoang MH, Lee HJ, Kim N, Lee D, Hwang KY, Hwang BY, Choi DW, Lee SJ. Ursolic acid is a PPAR-α agonist that regulates hepatic lipid metabolism. Bioorg Med Chem Lett. 2011; 21:5876–5880. PMID:
21855333.
47. Jia Y, Kim S, Kim J, Kim B, Wu C, Lee JH, Jun HJ, Kim N, Lee D, Lee SJ. Ursolic acid improves lipid and glucose metabolism in high-fat-fed C57BL/6J mice by activating peroxisome proliferatoractivated receptor alpha and hepatic autophagy. Mol Nutr Food Res. 2015; 59:344–354. PMID:
25418615.
48. Jang SM, Yee ST, Choi J, Choi MS, Do GM, Jeon SM, Yeo J, Kim MJ, Seo KI, Lee MK. Ursolic acid enhances the cellular immune system and pancreatic beta-cell function in streptozotocin-induced diabetic mice fed a high-fat diet. Int Immunopharmacol. 2009; 9:113–119. PMID:
19013541.
49. The World Health Organization MONICA Project (monitoring trends and determinants in cardiovascular disease): a major international collaboration. WHO MONICA Project Principal Investigators. J Clin Epidemiol. 1988; 41:105–114. PMID:
3335877.
50. Ness AR, Powles JW. Fruit and vegetables, and cardiovascular disease: a review. Int J Epidemiol. 1997; 26:1–13. PMID:
9126498.
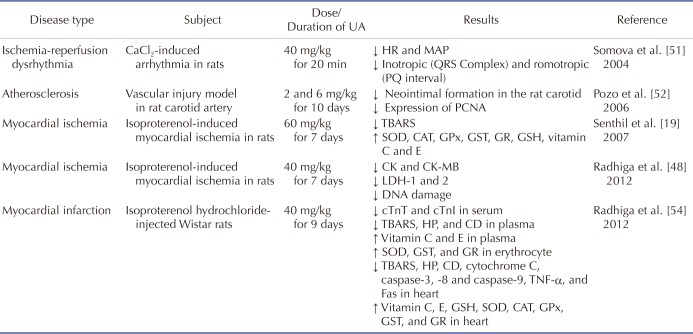
51. Somova LI, Shode FO, Mipando M. Cardiotonic and antidysrhythmic effects of oleanolic and ursolic acids, methyl maslinate and uvaol. Phytomedicine. 2004; 11:121–129. PMID:
15070161.
52. Pozo M, Castilla V, Gutierrez C, de Nicolás R, Egido J, González-Cabrero J. Ursolic acid inhibits neointima formation in the rat carotid artery injury model. Atherosclerosis. 2006; 184:53–62. PMID:
15922347.
53. Radhiga T, Rajamanickam C, Senthil S, Pugalendi KV. Effect of ursolic acid on cardiac marker enzymes, lipid profile and macroscopic enzyme mapping assay in isoproterenol-induced myocardial ischemic rats. Food Chem Toxicol. 2012; 50:3971–3977. PMID:
22898613.
54. Radhiga T, Rajamanickam C, Sundaresan A, Ezhumalai M, Pugalendi KV. Effect of ursolic acid treatment on apoptosis and DNA damage in isoproterenol-induced myocardial infarction. Biochimie. 2012; 94:1135–1142. PMID:
22289617.
55. Saravanan R, Pugalendi V. Impact of ursolic acid on chronic ethanol-induced oxidative stress in the rat heart. Pharmacol Rep. 2006; 58:41–47.
56. Lv YY, Jin Y, Han GZ, Liu YX, Wu T, Liu P, Zhou Q, Liu KX, Sun HJ. Ursolic acid suppresses IL-6 induced C-reactive protein expression in HepG2 and protects HUVECs from injury induced by CRP. Eur J Pharm Sci. 2012; 45:190–194. PMID:
22108347.
57. Bondy SC1, LeBel CP. The relationship between excitotoxicity and oxidative stress in the central nervous system. Free Radic Biol Med. 1993; 14:633–642. PMID:
8325535.
58. Castegna A, Aksenov M, Aksenova M, Thongboonkerd V, Klein JB, Pierce WM, Booze R, Markesbery WR, Butterfield DA. Proteomic identification of oxidatively modified proteins in Alzheimer's disease brain. Part I: creatine kinase BB, glutamine synthase, and ubiquitin carboxy-terminal hydrolase L-1. Free Radic Biol Med. 2002; 33:562–571. PMID:
12160938.
59. De Iuliis A, Grigoletto J, Recchia A, Giusti P, Arslan P. A proteomic approach in the study of an animal model of Parkinson's disease. Clin Chim Acta. 2005; 357:202–209. PMID:
15946658.
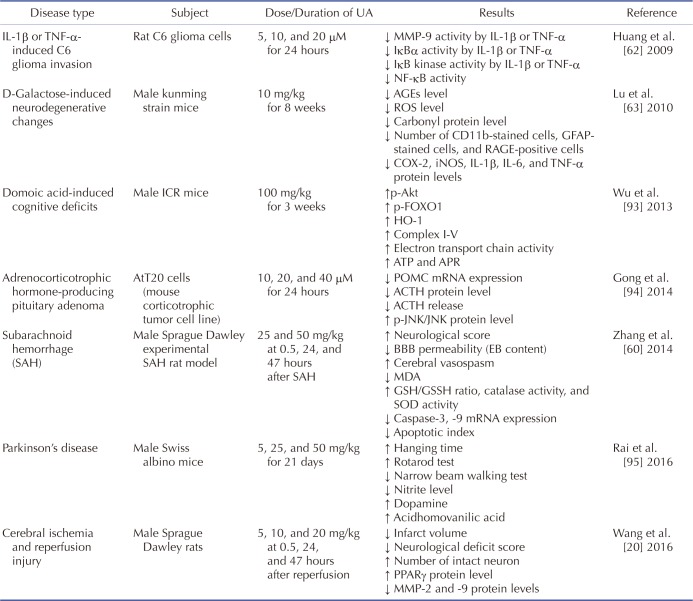
60. Zhang T, Su J, Wang K, Zhu T, Li X. Ursolic acid reduces oxidative stress to alleviate early brain injury following experimental subarachnoid hemorrhage. Neurosci Lett. 2014; 579:12–17. PMID:
25026072.
61. Shih YH, Chein YC, Wang JY, Fu YS. Ursolic acid protects hippocampal neurons against kainate-induced excitotoxicity in rats. Neurosci Lett. 2004; 362:136–140. PMID:
15193771.
62. Huang HC, Huang CY, Lin-Shiau SY, Lin JK. Ursolic acid inhibits IL-1beta or TNF-alpha-induced C6 glioma invasion through suppressing the association ZIP/p62 with PKC-zeta and downregulating the MMP-9 expression. Mol Carcinog. 2009; 48:517–531. PMID:
18973186.
63. Lu J, Wu DM, Zheng YL, Hu B, Zhang ZF, Ye Q, Liu CM, Shan Q, Wang YJ. Ursolic acid attenuates D-galactose-induced inflammatory response in mouse prefrontal cortex through inhibiting AGEs/RAGE/NF-κB pathway activation. Cereb Cortex. 2010; 20:2540–2548. PMID:
20133359.
64. Lu J, Zheng YL, Wu DM, Luo L, Sun DX, Shan Q. Ursolic acid ameliorates cognition deficits and attenuates oxidative damage in the brain of senescent mice induced by D-galactose. Biochem Pharmacol. 2007; 74:1078–1090. PMID:
17692828.
65. Rao JS, Steck PA, Mohanam S, Stetler-Stevenson WG, Liotta LA, Sawaya R. Elevated levels of M(r) 92,000 type IV collagenase in human brain tumors. Cancer Res. 1993; 53(10 Suppl):2208–2211. PMID:
8485704.
66. Ma JQ, Ding J, Zhang L, Liu CM. Protective effects of ursolic acid in an experimental model of liver fibrosis through Nrf2/ARE pathway. Clin Res Hepatol Gastroenterol. 2015; 39:188–197. PMID:
25459994.
67. Yang LJ, Tang Q, Wu J, Chen Y, Zheng F, Dai Z, Hann SS. Inter-regulation of IGFBP1 and FOXO3a unveils novel mechanism in ursolic acid-inhibited growth of hepatocellular carcinoma cells. J Exp Clin Cancer Res. 2016; 35:59. PMID:
27036874.
68. Tailleux A, Wouters K, Staels B. Roles of PPARs in NAFLD: potential therapeutic targets. Biochim Biophys Acta. 2012; 1821:809–818. PMID:
22056763.
69. Reddy JK, Rao MS. Lipid metabolism and liver inflammation. II. Fatty liver disease and fatty acid oxidation. Am J Physiol Gastrointest Liver Physiol. 2006; 290:G852–G858. PMID:
16603729.
70. Gao HY, Li GY, Lou MM, Li XY, Wei XY, Wang JH. Hepatoprotective effect of Matrine salvianolic acid B salt on Carbon Tetrachloride-Induced Hepatic Fibrosis. J Inflamm (Lond). 2012; 9:16. PMID:
22559721.
71. Son HS, Kwon HY, Sohn EJ, Lee JH, Woo HJ, Yun M, Kim SH, Kim YC. Activation of AMP-activated protein kinase and phosphorylation of glycogen synthase kinase3 β mediate ursolic acid induced apoptosis in HepG2 liver cancer cells. Phytother Res. 2013; 27:1714–1722. PMID:
23325562.
72. DeFronzo RA, Tripathy D. Skeletal muscle insulin resistance is the primary defect in type 2 diabetes. Diabetes Care. 2009; 32(Suppl 2):S157–S163. PMID:
19875544.
73. Gardner DS, Rhodes P. Developmental origins of obesity: programming of food intake or physical activity? Adv Exp Med Biol. 2009; 646:83–93. PMID:
19536666.
74. Carter HN, Chen CC, Hood DA. Mitochondria, muscle health, and exercise with advancing age. Physiology (Bethesda). 2015; 30:208–223. PMID:
25933821.
75. Seo DY, Lee SR, Kim N, Ko KS, Rhee BD, Han J. Age-related changes in skeletal muscle mitochondria: the role of exercise. Integr Med Res. 2016; 5:182–186. PMID:
28462116.
76. Cederholm TE, Bauer JM, Boirie Y, Schneider SM, Sieber CC, Rolland Y. Toward a definition of sarcopenia. Clin Geriatr Med. 2011; 27:341–353. PMID:
21824551.
77. Cederholm T, Morley JE. Sarcopenia: the new definitions. Curr Opin Clin Nutr Metab Care. 2015; 18:1–4. PMID:
25207791.
78. Goodpaster BH, Carlson CL, Visser M, Kelley DE, Scherzinger A, Harris TB, Stamm E, Newman AB. Attenuation of skeletal muscle and strength in the elderly: the Health ABC Study. J Appl Physiol (1985). 2001; 90:2157–2165. PMID:
11356778.
79. Masanes F, Culla A, Navarro-Gonzalez M, Navarro-Lopez M, Sacanella E, Torres B, Lopez-Soto A. Prevalence of sarcopenia in healthy community-dwelling elderly in an urban area of Barcelona (Spain). J Nutr Health Aging. 2012; 16:184–187. PMID:
22323356.
80. Viana JU, Silva SL, Torres JL, Dias JM, Pereira LS, Dias RC. Influence of sarcopenia and functionality indicators on the frailty profile of community-dwelling elderly subjects: a cross-sectional study. Braz J Phys Ther. 2013; 17:373–381. PMID:
23970115.
81. Amigues I, Schott AM, Amine M, Gelas-Dore B, Veerabudun K, Paillaud E, Beauchet O, Rolland Y, Canouï Poitrine F, Bonnefoy M. Low skeletal muscle mass and risk of functional decline in elderly community-dwelling women: the prospective EPIDOS study. J Am Med Dir Assoc. 2013; 14:352–357. PMID:
23333310.
82. Bakhtiari N, Hosseinkhani S, Tashakor A, Hemmati R. Ursolic acid ameliorates aging-metabolic phenotype through promoting of skeletal muscle rejuvenation. Med Hypotheses. 2015; 85:1–6. PMID:
25976755.
83. Ebert SM, Dyle MC, Bullard SA, Dierdorff JM, Murry DJ, Fox DK, Bongers KS, Lira VA, Meyerholz DK, Talley JJ, Adams CM. Identification and small molecule inhibition of an activating transcription factor 4 (ATF4)-dependent pathway to age-related skeletal muscle weakness and atrophy. J Biol Chem. 2015; 290:25497–25511. PMID:
26338703.
84. Jeong JW, Shim JJ, Choi ID, Kim SH, Ra J, Ku HK, Lee DE, Kim TY, Jeung W, Lee JH, Lee KW, Huh CS, Sim JH, Ahn YT. Apple pomace extract improves endurance in exercise performance by increasing strength and weight of skeletal muscle. J Med Food. 2015; 18:1380–1386. PMID:
26331671.
85. Ogasawara R, Sato K, Higashida K, Nakazato K, Fujita S. Ursolic acid stimulates mTORC1 signaling after resistance exercise in rat skeletal muscle. Am J Physiol Endocrinol Metab. 2013; 305:E760–E765. PMID:
23900420.
86. Chen J, Wong HS, Leong PK, Leung HY, Chan WM, Ko KM. Ursolic acid induces mitochondrial biogenesis through the activation of AMPK and PGC-1 in C2C12 myotubes: a possible mechanism underlying its beneficial effect on exercise endurance. Food Funct. 2017; 8:2425–2436. PMID:
28675237.
87. Bang HS, Seo DY, Chung YM, Oh KM, Park JJ, Arturo F, Jeong SH, Kim N, Han J. Ursolic Acid-induced elevation of serum irisin augments muscle strength during resistance training in men. Korean J Physiol Pharmacol. 2014; 18:441–446. PMID:
25352765.
88. Bang HS, Seo DY, Chung YM, Kim DH, Lee SJ, Lee SR, Kwak HB, Kim TN, Kim M, Oh KM, Son YJ, Kim S, Han J. Ursolic acid supplementation decreases markers of skeletal muscle damage during resistance training in resistance-trained men: a pilot study. Korean J Physiol Pharmacol. 2017; 21:651–656. PMID:
29200908.
89. Cho YH, Lee SY, Kim CM, Kim ND, Choe S, Lee CH, Shin JH. Effect of loquat leaf extract on muscle strength, muscle mass, and muscle function in healthy adults: a randomized, double-blinded, and placebo-controlled trial. Evid Based Complement Alternat Med. 2016; 2016:4301621. PMID:
27999607.
90. Church DD, Schwarz NA, Spillane MB, McKinley-Barnard SK, Andre TL, Ramirez AJ, Willoughby DS. l-Leucine increases skeletal muscle IGF-1 but does not differentially increase Akt/mTORC1 signaling and serum IGF-1 compared to ursolic acid in response to resistance exercise in resistance-trained men. J Am Coll Nutr. 2016; 35:627–638. PMID:
27331824.
91. Kazmi I, Afzal M, Rahman S, Iqbal M, Imam F, Anwar F. Antiobesity potential of ursolic acid stearoyl glucoside by inhibiting pancreatic lipase. Eur J Pharmacol. 2013; 709:28–36. PMID:
23500199.
92. Zhang Y, Song C, Li H, Hou J, Li D. Ursolic acid prevents augmented peripheral inflammation and inflammatory hyperalgesia in high-fat diet-induced obese rats by restoring downregulated spinal PPARα. Mol Med Rep. 2016; 13:5309–5316. PMID:
27108888.
93. Wu DM, Lu J, Zhang YQ, Zheng YL, Hu B, Cheng W, Zhang ZF, Li MQ. Ursolic acid improves domoic acid-induced cognitive deficits in mice. Toxicol Appl Pharmacol. 2013; 271:127–136. PMID:
23707761.
94. Gong YY, Liu YY, Yu S, Zhu XN, Cao XP, Xiao HP. Ursolic acid suppresses growth and adrenocorticotrophic hormone secretion in AtT20 cells as a potential agent targeting adrenocorticotrophic hormone-producing pituitary adenoma. Mol Med Rep. 2014; 9:2533–2539. PMID:
24682498.
95. Rai SN, Yadav SK, Singh D, Singh SP. Ursolic acid attenuates oxidative stress in nigrostriatal tissue and improves neurobehavioral activity in MPTP-induced Parkinsonian mouse model. J Chem Neuroanat. 2016; 71:41–49. PMID:
26686287.


 PDF
PDF ePub
ePub Citation
Citation Print
Print



 XML Download
XML Download