

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Acepromazine, belonging to the phenothiazine derivative antipsychotics, was developed in 1950s and used to treat psychosis in human. Since acepromazine did not show any significant therapeutic effect for the treatment of human psychiatric disorders, it is not clinically used at present [1234]. However, acepromazine is one of the most widely used sedative in veterinary medicine [5]. It is often pretreated in dogs and cats before anesthesia and also treated when large animals such as, horses and impalas are transported or examined [678]. In addition, acepromazine intoxications have been also reported in human. An accidental ingestion of acepromazine in a pediatric patient has been reported [9]. It also has been reported that acepromazine intoxicaes from drowsiness to death in adults when it was used to attempt to suicide or in drug-facilitated crimes [101112131415].
The human ether-à-go-go related gene (hERG) encodes the rapid delayed rectifier potassium current (Ikr), involving the regulation of cardiac rhythm by the repolarization of the membrane potential to the resting state during the cardiac action potential [16]. Blocking of hERG channels leads severe cardiovascular adverse effects such as acquired long QT syndromes and then increases risk for torsade de pointes arrhythmias, and sudden death [1718]. The mammalian ether-à-go-go gene families belonging to hERG are highly conserved and have common characters [19]. Mouse ether-à-go-go related gene (mERG) B is expressed selectively in the heart and has a similar electrophysiological feature with hERG B, the human homolog of mERG B [20]. The phenothiazine antipsychotics (thioridazine, chlorpromazine) inhibit hERG channels, which contribute a critical role in arrhythmogenesis [21222324]. Even though, acepromazine has been often used in veterinary medicine and few human intoxication of acepromazine has been reported, the toxicological mechanisms of acepromazine in animals as well as in human have not been studied yet. Thus, we investigated whether the hERG potassium channel, a major target of arrthythmogenic drugs, was affected by acepromazine to evaluate the toxic potentials in human and animal.
Go to : 
METHODS
Cell culture
The hERG-HEK293 recombinant cell line (CYL3039, Millipore, Billerica, MA, USA) was used for electrophysiological recording, as previously reported in detail [25]. The cells were maintained in D-MEM/F-12 (Invitrogen, Grand Island, NY, USA) supplemented with 10% fetal bovine serum, 1% nonessential amino acid, and 400 µg/ml geneticin, according to the manufacturer’s directions. The cells were plated on cover glasses (12 mm diameter; Fisher Scientific, Pittsburgh, PA, USA) and placed in 35 mm culture dishes at least 24 hours prior to patch -clamp recordings.
Solutions and drugs
The external bath solution contained 140 NaCl, 5 KCl, 1 CaCl2, 1 MgCl2, 10 HEPES, and 10 glucose in mM, and was adjusted to pH 7.3 using NaOH. Osmolarity of the solution measured using a vapor pressure osmometer (Vapro 5520, Wescor, Logan, UT, USA) was 300~310 mOsm. The internal pipette solution contained 140 KCl, 1 CaCl2, 1 MgCl2, 10 HEPES, 10 EGTA in mM and was adjusted to pH 7.3 using KOH. The average osmolarity of internal solution was 290 mOsm. Acepromazine (Santa Cruz Biotechnology, Dallas, Texas, USA) was dissolved in dimethylsulfoxide (DMSO, Sigma, St. Louis, MO, USA) as a stock solution of 100 mM, and the stock solution was then diluted with the external solution to obtain the desired concentration. The concentration of DMSO in the final dilution was <0.1%, and this DMSO concentration had no effect on the hERG currents [25]. The θ-tubing glass pipette mounted on a piezoelectric translator (P-601 PiezoMove Z Acturator, Physik Instrumente, Karlsruhe, Germany) was used for fast drug application. Solutions were rapidly switched around the cell using a piezoelectric translator displacing the θ-tubing laterally to expose the cells to the drug-containing solution for a defined period of time, and then rapidly return to the drug-free solution. Solutions were delivered under gravity from reservoirs placed above the preparation, and the application timing was controlled by a perfusion valve control system (VC-8, Warner Instruments, Hamden, CT, USA).
Electrophysiology
For electrophysiological recording, the cover glasses containing adherent hERG-HEK293 recombinant cells were transferred to the recording chamber (RC-13, Warner Instruments) mounted on the stage of an inverted microscope (IX70, Olympus, Tokyo, Japan). Cells were continuously perfused with an external bath solution. hERG currents were recorded using a Multiclamp 700 B microelectrode amplifier and pClamp 10.1 software (Molecular Devices, Sunnyvale, CA, USA) in a whole-cell configuration of the patch-clamp technique at room temperature (22~24℃). Glass micropipettes were pulled from glass capillaries (PG10165-4, World Precision Instruments, Sarasota, FL, USA) using a programmable horizontal microelectrode puller (P-97, Sutter Instrument, Novato, CA, USA). The tip resistances of the patch pipettes were 2~4 MΩ when filled with the internal solution. The liquid junction potentials between pipette and bath solutions were in the range 3~5 mV, and zeroed before the gigaohm seal was formed. The current signals were filtered at 2 kHz, digitized at 10 kHz, and saved on a PC using DigiData 1322 and pClamp 10.1 software (Molecular Devices).
Statistical analysis
Data analysis was performed using pClamp 10.1 software (Molecular Device) and Origin 8.0 software (Origin Lab Corp., Northampton, MA, USA). The averaged data are presented as the mean±S.E.M. A paired Student's t-test was used for the statistical analysis of paired data. A one-way analysis of variance with Bonferroni's test was applied for the comparison of multiple groups. p<0.05 was considered statistically significant.
Go to :
RESULTS
Concentration-response relationships for blocking of hERG currents by acepromazine
Fig. 1A shows the representative hERG currents traces recorded in the absence and presence of acepromazine (0.1, 0.3, 1, 3, and 10 µM). The hERG currents were evoked with 4 sec depolarization to +20 mV from a holding potential of –80 mV followed by a repolarization to –50 mV for 6 sec. The voltage protocol was applied every 15 sec. The peak tail currents of the hERG at –50 mV were normalized to the control values and plotted for drug concentrations (Fig. 1B) to evaluate the effect of acpromazine on the activated states of hERG channels. Acepromazine inhibited the hERG tail currents at –50 mV in a concentration-dependent manner. At the concentrations of 0.1, 0.3, 1, 3, and 10 µM acepromazine, the tail currents reduced by 16.3±1.7, 26.3±2.1, 46.4±2.8, 70.5±2.7, and 89.3±0.8% (n=10), respectively. The concentration-response curve was fitted by using Hill equation, yielding an IC50 value of 1.5±0.1 µM and Hill coefficient of 1.1±0.1 (n=10).
 | Fig. 1Concentration-dependent inhibition of hERG currents by acepromazine.(A) hERG currents were elicited by a 4-sec depolarizing pulse to +20 mV from a holding potential of –80 mV and repolarization to –50 mV for 6 sec to measure the peak tail currents in the absence and presence of acepromazine. The dotted line marks zero current. (B) Normalized hERG tail currents (Itail) to the control were plotted as a function of acepromazine concentration. The experimental data were fitted with a Hill equation. Data were expressed as the means±S.E.M.
|
Voltage-dependent inhibition of hERG currents by acepromazine
To identify the current-voltage (I -V) relationships of the hERG currents, the channels were activated by applying 4 sec depolarizing step-pulses (Δ10 mV, –70 mV~+60 mV) from a holding potential of –80 mV in every 15 sec followed by a 6 sec repolarizing pulse to –50 mV in the absence and presence of 1 µM acepromazine, which was the near IC50 value for hERG currents based on the Fig. 1. The steady-state hERG currents (Iss) were measured at the end of the depolarizing step-pulses and the peak hERG tail currents (Itail) were measured during the repolarizing periods (Fig. 2A). Fig. 2B shows the average I-V relationships for Iss and Itail before and after the application of 1 µM acepromazine. Under the control conditions, Iss was activated at a membrane potential of approximately –50 mV and reached the maximum at –20 mV. Acepromazine significantly inhibited the Iss at the membrane potentials between –40 and +60 mV (Fig. 2B, left). To investigate the effects of acepromazine on the voltage dependency of hERG channel activation step, the Itail were analyzed and normalized data were plotted against the membrane potential with or without acepromazine (Fig. 2B, right). Acepromazine significantly inhibited the Itail at the membrane potentials between –40 and +60 mV. When the plots of normalized tail currents were fitted to a Boltzmann function, acepromazine shifted the half-activation membrane potential (V1/2) of activation curves to the hyperpolarizing direction (i.e., from –38.6±0.8 mV to –45.0±0.8 mV, n=6, p<0.001), and decreased the slope factor (k) from 5.6±0.2 to 4.6±0.2 (p<0.001). The voltage-dependent inhibition of the Itail by acepromazine increased sharply for voltages of channel activation ranging from 28.6±4.5% at –40 mV to 56.2±1.6% at –10 mV (Fig. 2C, n=6, p<0.01). However, the inhibitions of Itail by acepromazine were not significantly different in the range of 0~+60 mV, and the voltage range after the channels were fully activated.
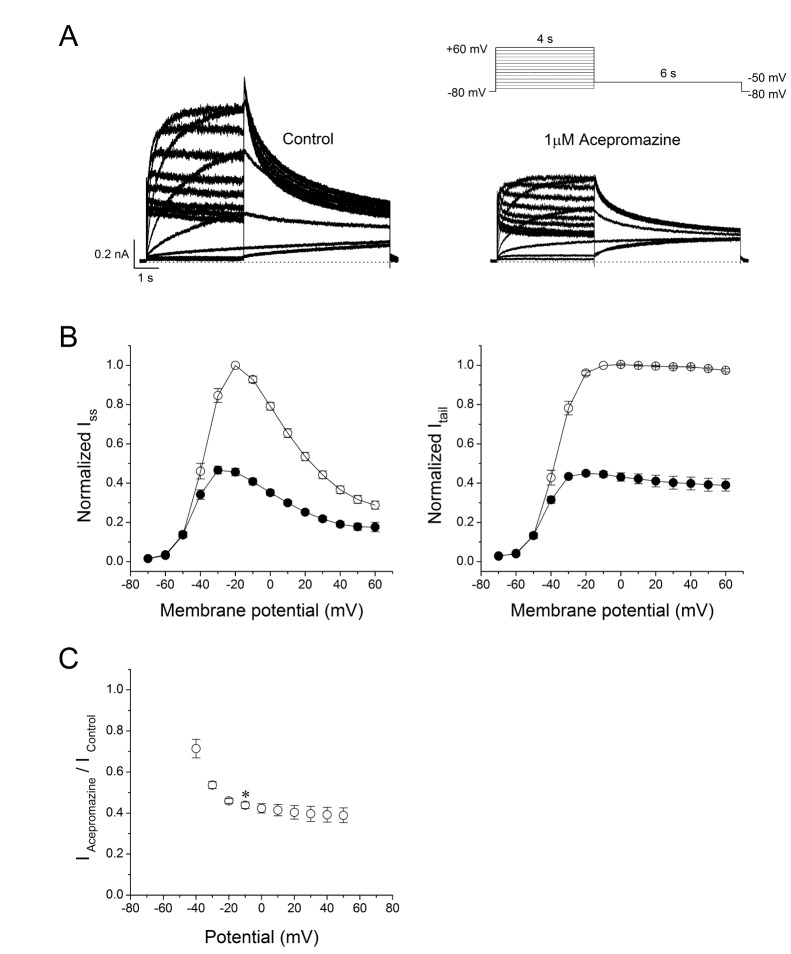 | Fig. 2Effect of acepromazine on current-voltage (I -V) relationships.(A) Whole-cell hERG currents were evoked by depolarizing pulses from –70 mV to +60 mV for 4 sec in steps of 10 mV every 15 sec from a holding potential of –80 mV and repolarization to –50 mV for 6 sec in the absence and presence of acepromazine (1 µM). (B) The I -V relationships of the steady-state (Iss) and peak tail (Itail) currents of the hERG channels under the control conditions (○) and after the application of acepromazine (●). The Iss and Itail at each voltage in the presence of acepromazine were normalized to those in the absence of acepromazine. Itail data were fitted to a Boltzmann function. (C) The voltage-dependent block of the Itail by acepromazine was expressed as a relative current (IAcepromazine/IControl). *Significant difference from data obtained at –10 mV when compared to –40 mV (p<0.01). Data are expressed as the means±S.E.M.
|
Interaction of acepromazine with both open and inactivated state of hERG channels
To investigate the effect of acepromazine with both the inactivated and open state of the hERG channels, a fast drug perfusion system was used [2526]. The hERG currents were elicited by a 5 sec depolarizing pre-pulse to +60 mV from a holding potential of –80 mV, followed by 5 sec of repolarization to –40 mV (Fig. 3A, n=9). Acepromazine (1 µM) was applied from the middle of depolarizing pulse to the end of repolarizing pulse, inhibiting the steady-state hERG currents by 27.9±1.3% at the end of the depolarizing pre-pulses (p<0.001) and the peak hERG tail currents by 35.0±1.5% immediately after repolarization to –40 mV (Fig. 3B, p<0.001). Thus, acepromazine preferentially inhibited the hERG channels in the open states than the inactivation.
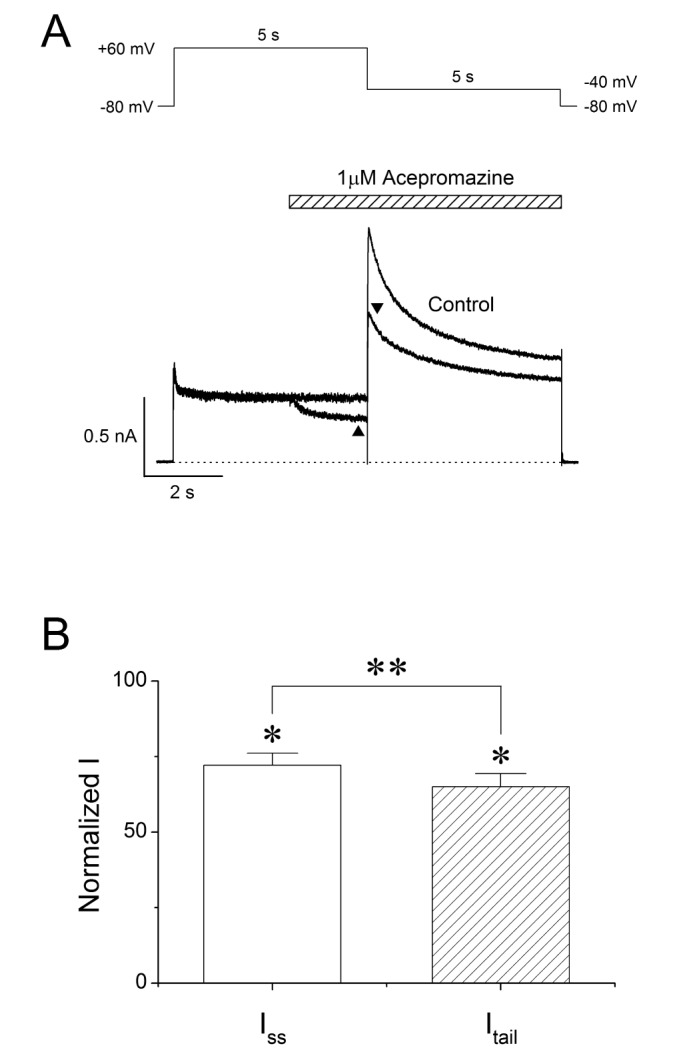 | Fig. 3Interaction of acepromazine with inactivated and open state of hERG channels. (A) The hERG currents were elicited by 5-sec depolarizing pulse to +60 mV from a holding potential of –80 mV, followed by a repolarizing pulse to –40 mV for 5 sec to induce peak tail currents. Acepromazine (1 µM) was rapidly applied during the depolarizing pulse and continued to the repolarizing pulse as indicated by the hatched bar. The currents were measured at the end of depolarization pulse (▲, Iss) and at the time of peak tail currents (▼, Itail), during the drug application. The dotted line marks zero current. (B) Averaged data showed that the inhibition of Iss and Itail by acepromazine. *p<0.001 when compared to control. **p<0.001 when compared between Iss and Itail. Data are expressed as the means±S.E.M.
|
Open channel block of the hERG currents by acepromazine
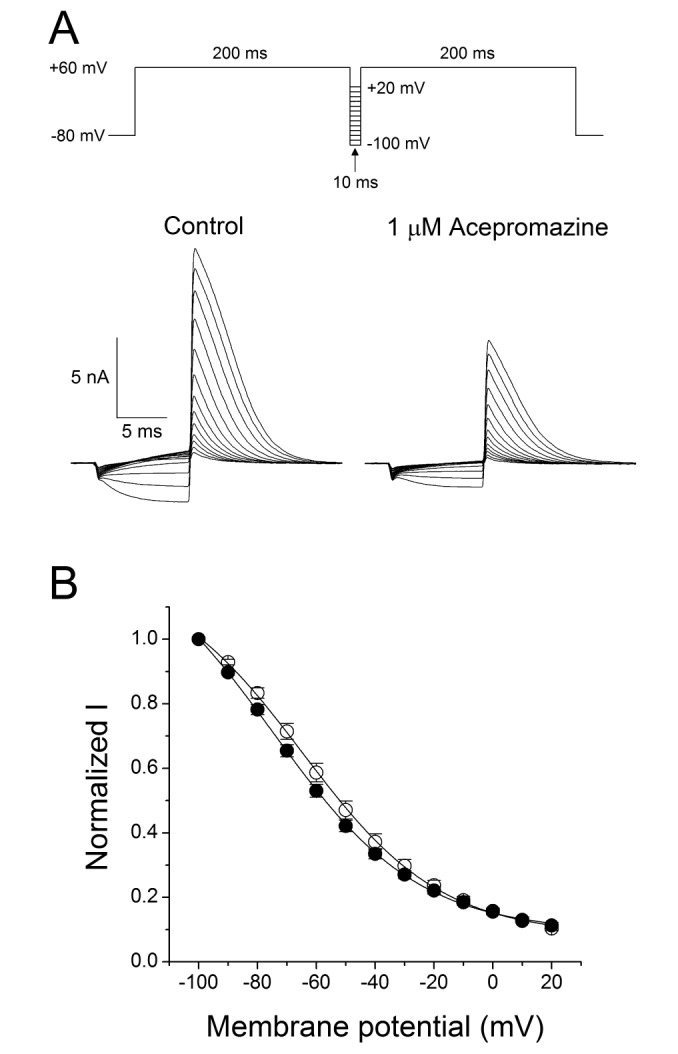
Fast drug application system was also used for further investigation of the acepromazine effect on the open states of hERG channels. The hERG currents were elicited by a 1sec depolarizing pre-pulse to +60 mV, rapidly inactivating the hERG channels followed by 10 sec of repolarization to –40 mV, and this pulse protocol induced a fast recovery of hERG channels from inactivation and remained hERG channels to the open states. Acepromazine was applied and removed quickly during the peak hERG tail currents for 5 sec. Fig. 4A shows a rapid and reversible inhibition of hERG tail currents by fast application of acepromazine in a concentration-dependent manner. The hERG tail currents was reduced by 4.5±0.7, 11.5±0.6, 27.2±1.2, 55.2±1.7, and 78.4±1.7% at 0.1, 0.3, 1, 3, and 10 µM of acepromazine, respectively (Fig. 4B, n=12). Based on this experiment, the calculated IC50 value for the open states of hERG channels was 2.4±0.2 µM with Hill coefficient of 1.2±0.1. This IC50 value was higher than the IC50 value for the activated states of hERG channels in the above experiment (Fig. 1B, IC50=1.5 µM) with almost the same value of Hill coefficient (1.1 vs. 1.2). This difference in the IC50 values was most likely explained by a higher affinity of acepromazine for the activated state of hERG channels than the open state.
 | Fig. 4Concentration-dependent effect of acepromazine on open state hERG channels.(A) The hERG currents were elicited by a 1-sec depolarizing pulse to +60 mV from a holding potential of –80 mV, followed by a repolarizing pulse to –40 mV for 10 sec to evoke peak tail currents. 1 sec after the start of repolarizing pulse, acepromazine was rapidly applied for 5 sec and then removed during the repolarizing pulse by fast drug application system. The bar indicates the time for the application of acepromazine. (B) Graph showed a concentration-response relationship for an open channel block of the hERG current by acepromazine. The normalized currents at the end of the drug application (▲) were plotted for the concentrations of acepromazine. The data were fitted using a Hill equation. Data were expressed as the means±S.E.M.
|
Effect of acepromazine on steady-state inactivation of hERG channels
To study the effect of acepromazine on the steady-state inactivation of hERG channels, the double-pulse protocol with steps of inter-stimulus repolarizing pulse was used (Fig. 5A). The cells were depolarized to +60 mV for 200 msec to induce inactivation and hyperpolarized from –100 mV to +20 mV in 10-mV voltage steps for 10 msec. And then a second depolarizing pulse of +60 mV was applied for 200 msec. Peak current amplitudes during the second depolarizing pulses were analyzed and normalized data to the maximum values were plotted to evaluate the steady-state inactivation of hERG channels. The acquired data under the control condition and 1 µM acepromazine were fitted to Boltzmann equation (Fig. 5B). The half-inactivation voltage (V1/2) and slope factor (k) were –64.2±3.5 mV and 22.8±0.4 in controls (n=7), and –74.7±3.1 mV and 24.5±0.4 in the presence of 1 µM acepromazine (n=7). The steady-state inactivation curve shifted to the hyperpolarized direction by acepromazine (p<0.001). However, the slope factor did not change significantly in the presence of acepromazine.
 | Fig. 5Effect of acepromazine on steady-state inactivation of hERG channels.(A) Representative current traces showed the steady-state inactivation under the control conditions and in the presence of 1 µM acepromazine, which were evoked by a double-pulse protocol with steps of inter-stimulus repolarizing pulses. After the first 200-msec depolarizing pulse of +60 mV inducing hERG channel inactivation, 10 msec inter-stimulus step pulses from –100 to +20 mV, with a 10 mV increment were applied, followed by a second depolarizing pulse of +60 mV. (B) Normalized steady-state inactivation curves under control conditions (○) and in the presence of 1 µM acepromazine (●). Solid lines represent fitting with Boltzmann function. Data are expressed as means±S.E.M.
|
Go to :
DISCUSSION
The effect of acepromazine on the hERG potassium channels expressed in HEK293 cells was investigated using the patch-clamp technique. The results of our study can be summarized as follows: 1) acepromazine inhibited the hERG channels in a concentration-dependent manner; 2) the drug also blocked hERG channels voltage dependently between –40 mV and +60 mV; 3) acepromazine inhibited the hERG currents by acting on both the inactivated (during the depolarization) and open (during the repolarization) states of the channels; 4) the result of fast drug application study was consistent with the open-channel block mechanism of hERG channel by acepromazine. These results suggest that acepromazine directly inhibits the hERG potassium channels, responsible for the repolarization of action potential in cardiac tissues, preferentially by binding to the open and inactivated states of ion channels.
Even though, the primary pharmacological mechanisms of neurobehavioral effects of phenothiazine antipsychotics, including acepromazine could be attributed to their potent antagonism of dopamine receptors [272829], these drugs also caused many central and peripheral adverse reactions by acting on the dopamine receptors (on-target adverse effects) and by nonspecific actions on other receptor types and ion channels (off-target adverse effects) [303132]. One of serious morbidity after the ingestion of the phenothiazine antipsychotics usually resulted from cardiotoxicity including hypotension, conduction delay and ventricular arrhythmia [33]. Especially, it has been reported that thioridazine and chlorpromazine directly inhibited hERG channels, through which these drugs could induce long QT syndrome [21222324]. Taken together with several reports showing that hERG currents inhibition is an important arrhythmogenic mechanism of acquired long QT syndrome and torsade de pointes [1718], our results suggest that acepromazine can cause arrhythmia possibly through the inhibition of hERG potassium channels as an off-target adverse effect.
cDNAs cloned from erg mRNA in cardiac tissue of the rat, guinea pig, rabbit, dog, and human showed a highly homologues. The dog and rabbit cDNAs were 99% identical to the human sequence and the rat and guinea pig’ were 96% identical to the human sequences [34]. Drugs reported to induce QT prolongation or torsade de pointes in human also prolonged QT intervals with 92% accuracy in canine Purkinje fiber model, an in vivo model for acquired long QT syndrome and drug-induced arrhythmogenesis [35]. Furthermore, canines have an important role in the preclinical assessment of QT interval prolongation in drug toxicity test [36]. A study assessing the potential neurobehavioral and cardiovascular toxicity of candidate drugs in a canine model observed that the dog’ heart rate was increased by acepromazine treatment [37]. Therefore, the drug-induced cardiac toxicity, especially related to long QT syndrome, seems to shares common molecular targets in both human and some other species such as dog, and rabbit.
It has been also reported that heart rates were increased in the two human intoxication cases of acepromazine, even though one of them was an overdose of acepromazine and etorphine combination [1014]. These results suggest that acepromazine overdoses can induce certain forms of adverse cardiovascular effects, including arrhythmias. However, there were still no reports showing that acepromazine induced the arrhythmias in human overdose cases and even veterinary uses.
The pharmacokinetic or toxicokinetic data of acepromazine in human have been not well documented. For veterinary use, the potency of acepromazine is species dependent: from 0.5 mg/kg in dogs to 5 mg/kg in horse, and the maximum plasma concentration (Cmax) of acepromazine was ~0.19 µM when administered at a dose of 0.15 mg/kg [38] and the Cmax of ~0.18 µM when administered at a dose of 0.1 mg/kg [39] in horses. In a human toxicology case, after 8 hr of 950 mg acepromazine ingestion, the plasma acepromazine level was 0.19 µM [10]. In our study, acepromazine inhibited hERG channel with an IC50 value of 1.5 µM, which was about ten times higher than the Cmax based on the sedative uses of acepromazine in the animals. These results suggest that acepromazine inhibits the hERG channels with relatively low potency. Chlorpromazine, a closely related structural analog of acepromazine, showed a higher IC50 value (21.6 µM) with a relative potential of long QT syndrome, even though its therapeutic plasma concentrations range (61.4~260 nM) were much lower than the IC50 [2324]. Because phenothiazines have a high lipophilicity and high affinity to lipids in the tissues, the tissue concentration of acepromazine was expected to be higher than plasma level [40]. Together with these reports, our results suggest that if acepromazine accumulates in cardiac tissue with high affinity, it has an arrhythmogenic potential through the inhibition of hERG potassium channels in cardiac tissues even at lower plasma concentration level.
Acepromazine has been widely used as a sedative in combination with various anesthetics or analgesics in veterinary medicine [541]. Moreover, many sedatives and anesthetics have been known to induce long QT syndrome though the inhibition of hERG potassium channels in human [42] and animals [43]. Even though acepromazine has a relatively higher IC50 for hERG channel inhibition, if it used in combination with drugs with potential of prolongation of QT intervals through the hERG channel inhibition, such as sedatives and anesthetics, it might be dangerous to human and animals through a synergistic drug interaction. This interpretation could be support by the reports that acepromazine significantly increased heart rate and blood pressure when it combined with halothane, ketamine, and butorphanol [4445]. These synergistic effects of hERG channel inhibition through the drug interaction could be another explanation of acepromazine possibly inducing long QT syndrome even at higher IC50 in our study.
This study shows that hERG currents were inhibited by acepromazine. In consistent with that hERG channels have high homology with other mammalian species, these results suggest that acepromazine have a potentially arrhythmogenic action through the inhibition of hERG potassium channels in cardiac tissues and the prolongation of QT intervals.
Go to :
 XML Download
XML Download