

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
The activation of synaptic and extrasynaptic γ-aminobutyric acid A receptors (GABAARs) generates phasic and tonic forms of inhibition (tonic GABAA current, Itonic), respectively [12], and has a profound influence on the hippocampal neural circuitry. Itonic is particularly interesting in the context of different ages because extrasynaptic GABAAR signaling is implicated in brain physiology and rage of pathophysiologies [34567]. Changes in extracellular GABA concentrations alter the relative contribution of specific GABAARs to Itonic as different receptor populations are recruited [8]. GABAARs containing the α5 subunit (α5-GABAARs) contribute to Itonic when the ambient GABA concentration increases, while at low ambient GABA concentrations the activation of δ subunit-containing receptors predominates [9]. In dentate gyrus granule cells (DGGCs), Itonic increases during initial postnatal maturation [1011], and further increases as adolescents mature into adulthood [12]. The age-dependent increase of Itonic in DGGCs may mirror the increased expression of δ-GABAARs in adults [13], which raises the question of whether and how a developmental change in α5-GABAARs alter Itonic at different ages.
GABA transporters (GATs) are members of a family of Na+-dependent neurotransmitter reuptake proteins. To date, four different GATs (GAT-1, GAT-2, GAT-3, and Betain/GABA transporter type 1) have been described in rat brain. Of these, GAT-1 is a primary neuronal GAT, while GAT-3 is commonly associated with glial cells [14]. Accordingly these two GAT subtypes are responsible for controlling extracellular GABA released from vesicular and non-vesicular sources, respectively [15]. In the hippocampus, GAT-1 predominantly determines the GABA concentration surrounding neurons, while GAT-3 activity is apparent with increased extracellular GABA concentration, especially when GAT-1 is blocked [16]. However, GAT-1 expression is low at early postnatal age, with GAT-3 expression dominating in that period [17]. Overall, it remains unknown whether and how the interaction between GAT-1 and GAT-3 modulates Itonic during postnatal brain maturation. In this study, we investigated the combined role of GAT-1 and GAT-3 in Itonic regulation in DGGCs at different ages; the results suggested that Itonic mirrored the changes in expression of extrasynaptic GABAARs activated by elevated extracellular GABA, according to the interrelationship between neuronal and glial GATs.
METHODS
Experimental animals
Male Sprague-Dawly rats purchased from Samtako Bio (Kyung Gi-Do, Korea) were housed under a 12/12 h light/dark schedule with free access to food and water until used. Animals were grouped by postnatal day (P), as follows: infantile (P7–9 and P14–16), juvenile (P21–23 and P28–30), adolescence (P36–37), and young adulthood (P42–44 and P49–51). Brains were rapidly extracted for electrophysiological recordings or Western blotting from animals anesthetized with ketamine and xylazine (80 mg/kg and 12 mg/kg, i.p., respectively). Animals in early infantile stage (P7–9) were euthanized by decapitation without anesthesia. All animal experimentation was conducted in compliance with the policies of Chungnam National University regarding the use and care of animals.
Electrophysiological recordings and data analysis
Patch-clamp recordings were obtained in acutely prepared coronal hippocampal slices from male rats, as described previously [618]. Briefly, slices were perfused with artificial cerebrospinal fluid (aCSF; in mM: NaCl 126, KCl 2.5, MgSO4 1, NaHCO3 26, NaH2PO4 1.25, glucose 20, ascorbic acid 0.4, CaCl2 1, pyruvic acid 2; pH 7.3~7.4; saturated with 95%O2–5%CO2) at a ~3 ml/min flow. Recordings were obtained at 32℃ using an Axopatch 200B amplifier (Axon Instruments, Foster City, CA). The series resistance was motored throughout the experiments. Neurons localized in the outer half of the granule cell layer were selected to minimize the effects of neurogenesis [19]. Patch pipettes were filled with a high Cl− containing solution (in mM): KCl 140, HEPES 10, Mg2+ATP 5, MgCl2 0.9, and EGTA 10. Current output was filtered at 2 kHz and digitized at 10 kHz (Digidata 1322A, pClamp 9 software, Axon Instruments). Itonic was defined as the difference between the holding current (Iholding) before and after application of the GABAA receptor blocker bicuculline (20 µM). Drugs were added to the perfusing aCSF solution at known concentrations. All drugs except NO-711 (Tocris, UK) were purchased from Sigma-Aldrich (St. Louis, MO, USA).
Western blotting
All proteins from the dissected dentate gyrus (DGs) were lysed with 1× passive lysis buffer (Cell Signaling Technology, Danvers, MA, USA) and quantified using a Coomassie Protein assay kit (Bio-Rad, Hercules, CA, USA). Approximately 50 µg of protein was electrophoresed on a 10% sodium dodecyl sulfate polyacrylamide gel (SDS–PAGE) and transferred onto nitrocellulose membranes. The blots were blocked with 1× Tris buffered saline (TBS)-Tween 20 containing 3% bovine serum albumin (BSA) +2% heparan sulfate (HS) for 1 h at room temperature (5% TTBS; Gibco, USA). The blots were then incubated at 4℃ with primary antibodies against GABAAR α5 subunit, GAT-1, and GAT-3 (1:1,000; Millipore, USA) in 5% TTBS, respectively. The next day, the blots were incubated with a horseradish peroxidase-conjugated goat anti-rabbit secondary antibody (1:2,000; Santa Cruz Biotechnology, USA). An enhanced chemiluminescence detection kit (ECL; Pierce, USA) was used to visualize antibody binding, and the intensity of the bands was measured using Image J software 1.42q (NIH, USA).
RESULTS
Electrophysiological recordings were obtained from a total of 114 DGGCs. In addition to blocking synaptic transmission, the GABAAR antagonist, bicuculline (BIC, 20 µM), induced a variable outward shift in the holding current (Iholding) in DGGCs.
Increased expression of GAT-1 and GAT-3 in preadolescence
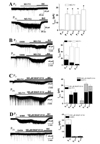
Altered expression of neuronal and glial GATs could contribute to Itonic changes by altering the extracellular GABA concentration at different ages. To understand the age-dependent changes in GAT expression, we directly compared the expression of GAT-1 and GAT-3 in the DGs at different ages. Western blot analysis showed that the expression of GAT-1 and GAT-3 in the DGs gradually increased at P7, P14, and P21, and thereafter stabilized at P28, P35, and P42 (Fig. 1).
Effect of GAT-1 and GAT-3 blockers on Itonic according to age
Changes in the expression and/or relative contribution of GAT-1 and/or GAT-3 may cause Itonic changes at different ages. To discriminate the functional role of neuronal and glial GATs in Itonic, we measured and compared the Itonic of DGGCs in the presence of selective GAT-1 and GAT-3 blockers at different ages.
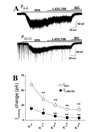
Bath application of NO-711 (5 µM), a selective GAT-1 blocker [20], failed to induce consistent changes in Iholding at P6–8, while it induced a similar inward shift in Iholding (INO-711) at P13–15 (21.8±4.4 pA, n=7), P20–22 (20.8±6.1 pA, n=6), P27–29 (21.4±3.6 pA, n=5), and P34–36 (27.3±3.4 pA, n=7) (Fig. 2A). In a subset of experiments, we measured INO-711 in the presence of 1 µM GABA to confirm that GAT-1 activity facilitates Itonic at the early infantile age. GABA (1 µM)-induced Iholding shift were decreased with age (P6–8, 16.3±4.2 pA, P20–22, 7.6±1.7 and P34–36, 3.3±1.2 pA, n=5~6). Additional application of NO-711 (GABA+NO-711) caused a significant inward shift in Iholding even at P6–8, which was still much smaller than at P20–22 and P34–36 (p<0.01 in both cases; Fig. 2B).
To investigate the functional significance of changes in GAT-3 expression, a selective GAT-3 blocker, SNAP-5114, was used [14]. Because SNAP-5114 (100 and 300 µM) alone failed to elicit changes in Iholding, we sequentially applied NO-711 and NO-711+SNAP-5114. In the presence of NO-711, 300 µM SNAP-5114 induced a significant inward shift in Iholding (ISNAP-5114) at P6–8, P20–22, and P34–36. ISNAP-5114 was significantly smaller at P6–8 than at P20–22, and P34–36 (p<0.05 in both cases; Fig. 2C). Even in the presence of NO-711, 100 µM SNAP-5114 did not induce significant changes in Iholding at all tested age groups (Fig. 2C). Thus, in a subset of experiments, we assessed ISNAP-5114 in the presence of 1 µM GABA at different ages. GABA induced significant Iholding changes at P6–9 (22.6±3.5 pA, n=6), while it induced minimal changes in Iholding at P20–22 (1.3±1.0 pA, n=5) and P34–36 (1.9±0.9 pA, n=5). An additional 100 µM SNAP-5114 efficiently induced an inward shift in Iholding at P6–9, while it failed to cause significant changes in Iholding at P20–22 and P34–36 in the presence of 1 µM GABA (Fig. 2D). These results support the idea that GAT3 activity is apparent with an increased extracellular GABA concentration in the hippocampus [16].
Nipecotic acid-induced Itonic in preadolescence
To investigate the combined role of GAT-1 and GAT-3 in developmental Itonic changes, we measured and compared the Itonic of DGGCs in the presence of the nonselective GAT blocker, nipecotic acid (NPA), at different ages.
In contrast to selective GAT-1 or GAT-3 blockade, NPA (100 µM) induced a significant inward shift in the Iholding of DGGCs (INPA), blocked by BIC, at all tested age groups. Interestingly, INPA gradually decreased during the infantile and juvenile periods (P6–8, P13–15, P20–22, and P27–29), and stabilized in adolescence and young adulthood (P34–36 and P41–43) (Fig. 3A and B).
We hypothesize that the age-dependent INPA change may mirror the functional maturation of GABAARs rather than the upregulation of GAT expression in preadolescence, and that the large INPA may be due to a high ambient GABA level. Therefore, we directly compared INPA inhibition of L-655,708, an inverse agonist at the benzodiazepine binding site of α5-GABAARs [21]. The bath application of L-655,708 (5 µM) efficiently blocked INPA at all tested ages (p<0.01 in all cases; Fig. 3A). Interestingly, in agreement with gradual INPA attenuation with age, L-655,708-sensitive INPA also gradually decreased during preadolescence (Fig. 3B). As a result, the portion of L-655,708-sensitive total INPA did not differ by age.
Age-dependent decease in the GABAAR α5 subunit in preadolescence
To understand the functional changes in GABAARs according to age, we directly compared Itonic activated by exogenous GABA (5 µM) and their sensitivity to L-655,708 at different ages.
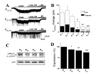
Itonic gradually decreases as infants mature into adolescence. The large INPA that characterized infantile periods (P6–8 and P13–15) gradually decreased at the juvenile (P20–22, and P27–29) and adolescent (P34–36) stages, and thereafter stabilized in young adults (P41–43) (Fig. 4A and B). The bath application of L-655,708 (5 µM) partially blocked Itonic in the presence of GABA at all tested ages (p<0.01 in all cases; Fig. 4A). In agreement with Itonic attenuation, L-655,708-sensitive Itonic gradually decreased in preadolescence (Fig. 4A and B). We observed a tendency for the portion of L-655,708-sensitive total Itonic to decrease with age, although this did not reach statistical significance.
In further experiments, we directly compared the expression of the GABAAR α5 subunit in DGs in the different age groups (Fig. 4C and D). Although we detected very low level of GABAARs α5 subunit immune reactivity at P7 and P14 in DGs, Western blot analysis showed that the expression of the GABAARs α5 subunit gradually decreased in the juvenile (P21 and P28) and adolescent (P35) periods, and thereafter stabilized in the young adults (P42). The degree of GABAARs α5 subunit expression was not further changed at P49 and P56 (data not shown).
DISCUSSION
The main findings of the present study were as follows: 1) agedependent increase in the INO-711 and ISNAP-5114 of DGGCs at infantile stages were partially consistent with the gradual increase in GAT-1 and GAT-3 expression in infantile and juvenile DGs; 2) the age-dependent decrease in INPA was compromised as GABAAR α5 subunit expression gradually decreased during preadolescence. Together, these findings suggest that, in addition to regulation of the ambient GABA concentration according to GAT activity, the change in age-dependent Itonic mirrored the altered expression and/or composition of extrasynaptic GABAARs during preadolescence.
Greater role of GAT-1 versus GAT-3 in regulating the Itonic of DGGCs
GATs embedded in axon terminal membranes and/or astrocyte plasma membranes regulate ambient GABA levels. In many neural systems, GAT-1 antagonists alone result in a smaller Itonic versus that observed when both GAT-1 and GAT-3 are blocked [2223]. This was explained by the involvement of both GAT-1 and GAT-3 transporters in regulating extracellular GABA concentrations around neurons. Similarly, INPA was much larger than INO-711 in infantile and juvenile DGGCs in the present study. Interpretation of this difference is complicated because, as opposed to NO-711, NPA is a GAT substrate. In addition, NPA could result in heteroexchange for GABA by GATs [24]. However, given that the concentrations of NPA and NO-711 used in this study were, respectively, about five and ten times that of the IC50 used for GAT-1 blockade [14], a simple interpretation could be that NPA blocked GAT-3 more efficiently than NO-711 during the infantile stage. However, our results showed that the GAT-1 blocker alone, and the GAT-1 blocker with additional application of SNAP-5114 (100 µM), resulted in a similar average Itonic of 20 pA in DGGCs after the late infantile periods; these results contradict the idea that GAT-3 actively contributed to the Itonic of DGGCs. In the present study, SNAP-5114 facilitated Itonic at the concentration of 300 µM, which was near to the IC50 for GAT-1 blockade [14], further confounding the role of GAT-3 in the Itonic of DGGCs. Combined with the fact that both GAT-1 and GAT-3 expression increased until the late juvenile periods, these results appear to support a major role of GAT-1 in the GABA uptake regulating Itonic in the adult hippocampus [16]. In general, our results suggest that GAT-1 and GAT-3 play a primary and adjunctive role, respectively, in regulating Itonic of DGGCs in preadolescece.
As with other neurotransmitter transporters, GATs can also act in reverse mode and thus release GABA from cells. Indeed, GABA can be secreted from cells by the reversed transport direction of GATs, particularly during early postnatal stages [2526]. Thus, it is possible that GABA release via reversed GAT activity is integral in maintaining GABA levels that activate Itonic [27] in the infantile and early juvenile stages. However, our finding that GAT-1 and GAT-3 blockers always enhanced the Itonic of DGGCs suggest that the two transporters operate synergistically to promote GABA uptake, which was seen at all tested ages in our experiments.
Role of α5-GABAARs in age-dependent INPA attenuation during preadolescence
Both the α5 and d subunit are key mediating components of the Itonic of DGGCs [28]. Alterations in GABA concentrations affect the relative contribution of specific GABAARs to Itonic as different receptor populations are recruited [8]. In the present study, BIC uncovered basal Itonic shown by the outward Iholding shift went over the initial level, especially when the inward shift in Iholding by exogenous GABA and GAT blockers was less than ~30 pA. α5–GABAARs contribute to Itonic when ambient GABA concentrations increase, while at low ambient GABA concentrations the activation of δ–GABAARs predominates [9]. In the present study, INPA was larger than INO-711 during preadolescence, which could be explained by an ambient GABA concentration sufficient to recruit additional α5–GABAARs in the presence of NPA, but not in NO-711. Our results showed that the portion of L-655,708-sensitive INPA ranged from ~27% to ~32%; this is consistent with previous findings showing that Itonic is mediated by α5–GABAARs and is responsible for ~29% of the total Itonic in DGGCs [28]. Thus, α5–GABAARs mediated INPA at all tested ages. Combined with the results whereby INPA and GABAAR α5 subunit expression gradually decreased with age, our results suggest that the age-dependent INPA decrease mirrors the functional decrease of α5–GABAARs rather than changes in GATs activity, during preadolescence.
However, there may be an as-yet undiscovered, non α5–containing GABAARs responsible for the large INPA observed during preadolescences. Indeed, in the present study, GABAAR α5 subunit immunoreactivity was not detectable in DGs at infantile stages. It is also notable that the small amplitude of INO-711 prevented us from comparing the sensitivity of INPA and that of INO-711 to L-655,708 in DGGCs. Regarding GABAARs activated in the presence of NPA, it is also noteworthy that NPA can directly activate GABAAR-like channels [29]. However, to the best of our knowledge, there is no information on the composition of GABAARs directly activated by NPA. However, it is still of interest that GABAAR α5 subunit expression gradually decreased with the postnatal development in various brain regions.
Functional consequences
Overall, our results showed that GATs blockades elevated ambient GABA level sufficiently to harmonize with α5-GABAARs, resulting in an age-dependent Itonic decrease in preadolescent brains. Combined with the fact that GABAAR α5 subunit expression in the hippocampus is closely related to learning and memory in young adults [3031], selective pharmacological modulators, such as α5-GABAAR selective inverse agonists, may be effective in increasing cognitive performance in memory disorders [32]. Future studies are warranted to elucidate the pathophysiology of α5-GABAARs generating Itonic combined with GAT blockades in the developing brains of preadolescents.

 XML Download
XML Download