

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
It is well known that excitation of high-threshold primary afferent nerve fibers, including thin myelinated (Aδ) or unmyelinated (C) fibers, triggers nociceptive pain. On other hand, selective excitation of low-threshold primary afferent nerve fibers, such as Aβ fibers, does not normally evoke pain. Mechanical allodynia is the perception of a painful sensation after the activation of low-threshold mechanoreceptors, which do not normally provoke pain in areas close to the lesion [1]. Impulses in Aβ fibers evoked by the activation of low-threshold mechanoreceptors can contribute to mechanical allodynia in animal pain models and in patients with pain [23]. Pretreatment with resiniferatoxin (RTX), which depletes capsaicin receptor protein (TRPV1) in primary afferent fibers, did not affect interleukin-1 beta (IL-1β)-induced mechanical allodynia [4]. Moreover, toll-like receptor 5-mediated selective Aβ-fiber blockade by co-application of flagellin and QX-314 suppresses mechanical allodynia in neuropathic pain models produced by chronic constriction injury or streptozotocin injection [5]. These results suggest that large myelinated Aβ fibers are the primary afferent fibers involved in mediating sensory processing of mechanical allodynia.
During inflammation or nerve injury, some large myelinated Aβ fibers may gain access to the nociceptive system and elicit pain sensation, although stimulation of primary afferent Aβ fibers does not induce hyperalgesia or allodynia under normal conditions. The dis-inhibition of local dorsal horn circuits following GABA inhibitory dysfunction is involved in the development of mechanical allodynia under inflammatory conditions [67]. This phenomenon leads to the unmasking of local inhibitory action of low-threshold afferents within nociceptive pathways. Although previous reports have demonstrated that large myelinated Aβ fibers elicit mechanical allodynia under pathological conditions [45], the underlying mechanisms are less clear. Moreover, there is no behavioral evidence for an excitatory link between low-threshold inputs and nociceptive neurons in the trigeminal system.
We investigated the central processing mechanisms of an excitatory link from a low-threshold input to nociceptive neurons under inflammatory pain conditions. For this purpose, we examined pro-nociceptive or paradoxical anti-nociceptive effects in naïve and IL-1β-treated rats, respectively, after intracisternal administration of bicuculline, which is a GABAA receptor antagonist. Bicuculline-induced paradoxical anti-nociceptive effects were examined after pretreatment with RTX, which depletes TRPV1 in primary afferent fibers. We also investigated the role of a sodium potassium chloride cotransporter (NKCC 1) in the development of IL-1β-induced mechanical allodynia after intracisternal injection of bumetanide, which is an NKCC 1 inhibitor. Finally, we confirmed the observed changes in GABAA receptor-mediated currents in medullary dorsal horn (MDH) neurons in IL-1β-treated rats.
Go to : 
METHODS
Animals and surgery
Experiments were performed on male Sprague-Dawley rats weighing 230~280 g. The animals were maintained in a temperature-controlled room (23±1℃) with a 12/12 hour light-dark cycle (lights on at 7:00 AM). Food and water were freely available. All procedures involving the use of animals were approved by the Institutional Animal Care and Use Committee of the School of Dentistry, Kyungpook National University (approved No. 2015-0053), and were carried out in accordance with the ethical guidelines for the investigation of experimental pain in conscious animals proposed by the International Association for the Study of Pain. All treatments were randomized during the experiments. All experimental procedures were performed in a blind manner.
Under anesthesia (a mixture 40 mg/kg ketamine and 4 mg/kg xylazine), a polyethylene tube (PE10, Clay Adams, BD, Parsippany, NJ) was implanted in the subcutaneous area of one vibrissa pad, which enabled the injection of IL-1β, as described previously [89]. For injection site consistency, a tube was implanted in the same vibrissa pad (3rd row, 5th column). The anesthetized rats were mounted individually on a stereotaxic frame (model 1404, David Kopf Instruments, Tujunga, CA) and the PE10 tube was implanted for the intracisternal injection as described previously [10111213]. The PE10 tube was inserted through a tiny hole made in the atlanto-occipital membrane and dura using a 27-gauge syringe needle. The tip of the cannula was placed at the level of the obex. The tube was then guided subcutaneously to the top of the skull and secured in place with a stainless steel screw and acrylic dental resin. As intracisternal catheterization may produce motor dysfunction, any animals displaying such effects or those with an incorrectly positioned catheter after intracisternal catheterization were excluded from further analysis. Pontamine sky blue dye was injected after behavioral tests to confirm placement of the intracisternal cannula and the extent of drug dissemination. The animals were given 72 hours to recover after surgery, as this time period is sufficient for complete recovery [1415].
General procedures for behavioral testing
Each rat was placed in a customized acrylic cylinder cage (height: 40~60 mm, length: 70~120 mm) for the evaluation of mechanical allodynia or thermal hyperalgesia. Each cage had a hole in the top so that the head could receive stimuli for the head withdrawal protocol. Each cage was placed in a darkened and noise-free room, and the animals were habituated for at least 30 minutes before starting the experiment. All behavioral tests were conducted between 1700 and 1800 h. The number of animal in each behavioral study group was eight.
Evaluation of mechanical allodynia
Withdrawal responses produced by ten successive trials of ramped air-puff pressure (4 seconds in duration, 10 second intervals) were examined as described previously [16171819]. The intensity of the air-puff pressure was controlled using a pneumatic pump (BH2 system, Harvard Apparatus). Air puffs were applied through a 26-gauge metal tube (length, 10 cm) located 1 cm from the skin at a 90° angle. The air-puff threshold was determined to be the air-puff pressure at which each rat responded in at least 50% of the trials. The cut-off pressure for the air puff was 40 psi, as described previously [202122]. Naïve rats did not respond to pressures below 40 psi.
Evaluation of thermal hyperalgesia
After applying radiant heat, head withdrawal latency was determined as described in previous studies [111623]. Heat stimulation was applied using an infrared thermal stimulator (Infrared Diode Laser, LVI-808-10; LVI tech, Seoul, Korea) at 11 W and 18.1 A. This intensity of thermal stimulation produced stable latencies of approximately 10-12 seconds at a 10 cm distance from the heat source to the vibrissa pad. Each rat received two stimuli and the inter-stimulus interval for each trial was at least 5 minutes. A cut-off time of 20 seconds was applied in these experiments to prevent possible tissue damage.
RTX treatment
To deplete TRPV1, some rats received a single intraperitoneal injection of RTX (200 µg/kg; Sigma, St. Louis, MO). A previous report indicates that systemic injection of RTX in rats damages TRPV1-expressing sensory neurons and induces a long-lasting impairment of thermal nociception, but does not affects mechanical allodynia [2425]. Moreover, RTX-treated animals show a marked reduction in TRPV1 transcription in the dorsal root ganglion, but not in the spinal cord, 7 days after intraperitoneal injection of RTX [26]. Thus, RTX is used as a tool to study the roles of nociceptive pathways involving non-TRPV1 afferent neurons and their modulation.
Brain stem slice preparation and gramicidin perforated patch recording
Rats (n=14 controls; n=9 IL-1β-treated) were anesthetized using isoflurane and then sacrificed by cervical dislocation 12 hours after subcutaneous injection of IL-1β (1 ng/10 µL). Brains were rapidly removed, and coronal brainstem slices (300~450 µm) containing the medullary dorsal horn (MDH) were prepared with a vibratome (Microm, Walldorf, Germany) in ice-cold artificial cerebrospinal fluid (ACSF) containing (in mM) 126 NaCl, 2.5 KCl, 2.4 CaCl2, 1.2 MgCl2, 11 D-glucose, 1.4 NaH2PO4, and 25 NaHCO3 (pH=7.4; bubbled with 95% O2 and 5% CO2). The slices were allowed to recover in oxygenated ACSF for at least 1 hour at room temperature. The slices were then transferred to the recording chamber, submerged, and superfused continuously with ACSF at a rate of 4~5 mL/minute. The slices were viewed using an upright microscope (BX51WI, Olympus, Tokyo, Japan) and Nomarski differential interference contrast optics.
Gramicidin perforated patch recordings were performed using an Axopatch 200B amplifier (Axon Instruments, Foster City, CA). For perforated-patch recordings, gramicidin (Molecular probes) was dissolved in dimethyl sulfoxide (Sigma, St. Louis, MO). The solution was diluted to a final concentration of 2.5~5 µg/mL in the pipette solution immediately before use and sonicated for 10 minute [27]. The pipette solution was passed through a disposable 0.22 µm filter and consisted of the following (in mM): 140 KCl, 1 CaCl2, 1 MgCl2, 10 N-2-hydroxyethylpiperazine-N'-2-ethanesulfonic acid, and 10 EGTA (pH=7.3 with KOH). The tip of the electrode was loaded with a small volume of gramicidin-free pipette solution and the electrode was back filled with the gramicidin-containing solution. The tip resistances of the recording electrodes ranged from 5~8 MΩ. Patch pipettes were pulled from thin-wall borosilicate glass-capillary tubing (PG52151-4, WPI, Sarasota, FL) using a Flaming Brown puller (P-97; Sutter Instruments Co., Novato, CA).
During the initial experiments, access resistance was monitored and experiments began when resistance was stabilized between 50 and 100 MΩ. Recordings began 15~20 minutes after gigaseal formation. As spontaneous rupture of the membrane was evident by a sudden over shooting of action potentials above 0 mV, data from spontaneously ruptured cells were not analyzed. Data were collected and analyzed with Clampex10.2 software (Axon Instruments, NY).
Experimental protocols
Effects of intracisternally injected bicuculline in naïve or IL-1β-treated rats
Bicuculline (20, 50, 100, or 200 ng/10 µL), a GABAA receptor antagonist, was administered intracisternally in naïve rats. Air-puff thresholds or head withdrawal latencies were examined 10, 20, 30, 45, 60, and 90 minutes after bicuculline injection. The present study also examined mechanical allodynia and thermal hyperalgesia in the IL-1β-treated rats after intracisternal administration of bicuculline. Bicuculline (50 ng/10 µL) was administered intracisternally 60 minutes after subcutaneous injection of IL-1β (1 ng/10 µL). Air-puff thresholds or head withdrawal latencies were examined 10, 20, 30, 45, 60, and 90 minutes after bicuculline injection.
Paradoxical anti-nociceptive effects of intracisternally injected bicuculline in RTX-treated rats
RTX (300 µg/kg) was administered intraperitoneally to exclude the role of TRPV1-dependent primary afferent fibers. IL-1β-induced mechanical allodynia was examined 2 days after injection of RTX. Intracisternal administration of 50 ng of bicuculline was performed 60 minutes after subcutaneous injection of 1 ng of IL-1β in RTX-treated rats. Air-puff thresholds were examined 10, 30, 60, and 90 minutes after bicuculline injection.
Evaluation of the role of NKCC1 in IL-1β-induced mechanical allodynia
Intracisternal administration of 50, 100, or 200 µg/10 µL of bumetanide, an NKCC1 inhibitor, was performed in naïve rats. We also intracisternally administered 50 or 100 µg of bumetanide 60 minutes after subcutaneous injection of 1 ng of IL-1β. Air-puff thresholds were examined 10, 20, 30, 40, 50, and 60 minutes after bicuculline injection.
Evaluation of the conversion of GABA-induced hyperpolarization to depolarization in IL-1β-treated rats
GABA (100 µM) and muscimol (3 µM), a specific GABAA receptor agonist, were applied in gramicidin perforated current clamp mode in order to compare their effects on the membranes of MDH neurons in IL-1β-treated rats. In the control group, we examined the effects of GABA (100 µM) and muscimol (3 µM) on the membranes of MDH neurons in saline-treated rats.
Drugs
IL-1β was purchased from R&D systems (Minneapolis, MN). Bicuculline, GABA, bumetanide and RTX were obtained from Sigma (St. Louis, MO). Muscimol was purchased from Tocris (UK). Gramicidin was purchased from Molecular Probes (ThermoFisher Scientific). IL-1β and bicuculline were dissolved in sterile saline, and bumetanide was dissolved in sterile saline containing 70% DMSO. Gramicidin was dissolved in 100% DMSO. RTX was dissolved in a mixture containing 10% ethanol and 10% Tween 80 in sterile saline.
Data analysis
Behavioral differences between groups were analyzed using a repeated measures ANOVA followed by Tukey honest significant difference post hoc analysis. The ration of depolarization induced by GABA or muscimol between the control and IL-1β-injected group was compared using a χ2-test. In all statistical comparisons, p<0.05 was used as the criterion for statistical significance. All data are presented as means±standard errors of the mean (SEMs).
Go to :
RESULTS
Effects of intracisternally injected bicuculline in naïve rats
Fig. 1 shows the changes in air-puff threshold and head withdrawal latencies after intracisternal administration of bicuculline. Intracisternal administration of the vehicle did not affect the air-puff thresholds. However, 20, 50, and 100 ng of bicuculline produced significant mechanical allodynia (F(3,28)=229.237, p<0.05, Fig. 1A). Intracisternal administration of 20 ng of bicuculline decreased air-puff thresholds significantly for 20 minutes after injection. The decreased air-puff thresholds produced by 50 and 100 ng of bicuculline persisted for 60 minutes after the injection. In contrast to the changes in the air-puff thresholds, only high dose bicuculline (200 ng) significantly decreased head withdrawal latencies after intracisternal administration (F(3,28)=3.373, p<0.05, Fig. 1B). Neither administration of the vehicle nor 50 or 100 ng of bicuculline affected head withdrawal latencies.
 | Fig. 1Effects of intracisternally administered bicuculline, a GABAA receptor antagonist, on air-puff thresholds and head withdrawal latency.(A) Treatment with vehicle did not affect air-puff thresholds. However, intracisternal administration of bicuculline (20, 50, or 100 ng) decreased air-puff thresholds significantly. (B) Neither vehicle nor low-dose bicuculline (50 ng) affected head withdrawal latencies. Intracisternal administration of bicuculline (100 or 200 ng) significantly decreased head withdrawal latency. *p<0.05, vehicle vs. bicuculline-treated group. Eight animals were used per group.
|
Effects of intracisternally injected bicuculline in IL-1β-treated rats
Fig. 2 shows the effects of intracisternal injection of bicuculline on IL-1β-induced mechanical allodynia and thermal hyperalgesia. Subcutaneous administration of 1 ng of IL-1β significantly decreased air-puff thresholds and IL-1β-induced mechanical allodynia persisted for 24 hours. Treatment with the vehicle 60 minutes after IL-1β injection did not affect IL-1β-induced mechanical allodynia. However, intracisternal administration of 50 ng of bicuculline reversed the reductions in the air-puff thresholds produced by IL-1β treatment when compare to the vehicle treatment (F(1,14)=115.488, p<0.05, Fig. 2A). The anti-allodynic effects of bicuculline persisted for 60 minutes after injection. Fig. 2B shows the effects of intracisternally injected bicuculline on IL-1β-induced thermal hyperalgesia. Intracisternal administration of 50 ng of bicuculline did not affect head withdrawal latencies in the IL-1β-treated group.
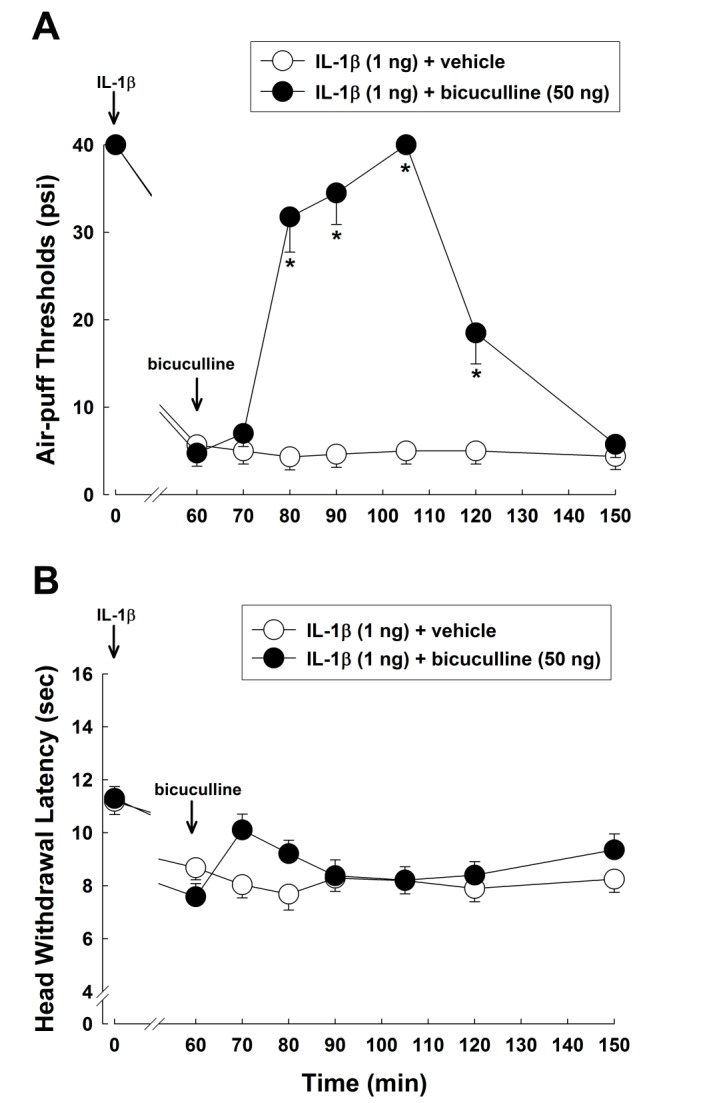 | Fig. 2Effects of intracisternally injected bicuculline on IL-1β-induced mechanical allodynia and thermal hyperalgesia.(A) Subcutaneous injection of IL-1β (1 ng) produced a significant decrease in air-puff thresholds. Intracisternal administration of bicuculline (50 ng) reversed the IL-1β-induced mechanical allodynia compared to vehicle treatment. (B) Subcutaneous injection of IL-1β (1 ng) significantly decreased head withdrawal latencies. However, intracisternal administration of bicuculline (50 ng) did not affect the IL-1β-induced thermal hyperalgesia. *p<0.05, vehicle vs. bicuculline-treated group. Eight animals were used per group.
|
Paradoxical anti-nociceptive effects of intracisternally injected bicuculline in RTX-treated rats
Fig. 3 illustrates the effects of pretreatment with RTX on bicuculline-induced paradoxical anti-allodynic effects in IL-1β-treated rats. In the vehicle-pretreated group, subcutaneous injection of IL-1β produced significant mechanical allodynia and intracisternal injection of bicuculline produced paradoxical anti-allodynic effects for 60 minutes following injection. Pretreatment with RTX did not affect the bicuculline-induced paradoxical anti-allodynic effects in IL-1β-treated rats.
 | Fig. 3Effects of intracisternally injected bicuculline on IL-1β-induced mechanical allodynia in RTX-treated animals.In the vehicle-pretreated group, subcutaneous injection of IL-1β decreased the air-puff threshold. Intracisternal administration of bicuculline produced paradoxical anti-allodynic effects. Pretreatment with RTX did not affect the bicuculline-induced paradoxical anti-allodynic effects. Eight animals were used per group.
|
Effects of intracisternally injected bumetanide on IL-1β-induced mechanical allodynia
IL-1β-induced mechanical allodynia was investigated after intracisternal injection of bumetanide (Fig. 4). In naïve rats, low doses of bumetanide (50 and 100 µg) did not affect air-puff thresholds, while high dose bumetanide (200 µg) decreased the air-puff thresholds (Fig. 4A). Therefore, low doses of bumetanide (50 and 100 µg) were used in the next experiment, as they did not produce mechanical allodynia in naïve rats. Intracisternal administration of low doses of bumetanide (50 and 100 µg) reversed the decrease in the air-puff thresholds produced by IL-1β injection (F(1,14)=211.281, p<0.05, Fig. 4B).
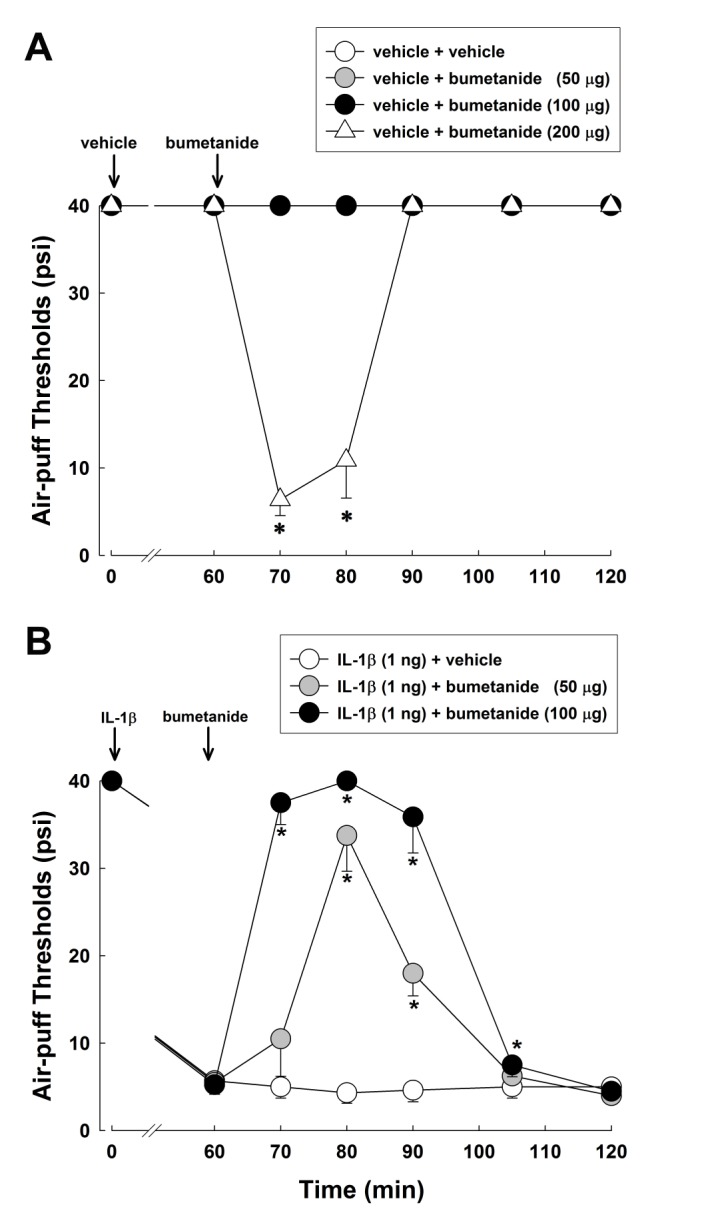 | Fig. 4Effects of intracisternally injected bumetanide, an NKCC1 inhibitor, on IL-1β-induced mechanical allodynia.(A) In naïve rats, low doses of bumetanide (50 or 100 µg) did not affect air-puff thresholds, while a high dose of bumetanide (200 µg) decreased air-puff thresholds. (B) Subcutaneous injection of IL-1β (1 ng) significantly decreased the air-puff thresholds. Intracisternal administration of low doses of bumetanide (50 or 100 µg) reversed the IL-1β-induced mechanical allodynia. *p<0.05, vehicle vs. bumetanide-treated group. Eight animals were used per group.
|
Conversion from GABA-induced hyperpolarization to depolarization in IL-1β-treated rats
Fig. 5A contains representative traces of GABA- or muscimol-induced membrane potential changes in the control group. In the control group, GABA (100 µM) induced membrane hyperpolarization in 10 (resting membrane potential [RMP]=−61.5±2.53 mV) of 18 MDH neurons tested (RMP=−62.1±2.11 mV). However, the other 8 neurons (RMP=−62.9±3.74 mV) had no response. Similarly, muscimol (3 µM) induced membrane hyperpolarization in 7 (RMP=−60.7±4.08 mV) of 15 MDH neurons tested (RMP=−61.5±2.64 mV), while the other 8 neurons (RMP=−62.0±3.69 mV) had no response. Membrane depolarization was not detected following GABA or muscimol addition to MDH neurons in the control group. Fig. 5B shows representative traces of GABA or muscimol-induced changes in membrane potential in the IL-1β-treated group. In the IL-1β-treated group, GABA (100 µM) induced membrane hyperpolarization in only 4 (RMP=−65.9±4.26 mV) of 17 MDH neurons (RMP=−63.0±2.17 mV), with no response observed in ten neurons (RMP=−63.6±2.30 mV). Interestingly, membrane depolarization was observed in 3 (RMP=−57.5±8.20 mV) of 17 neurons. Similarly, muscimol (3 µM) induced membrane hyperpolarization in 5 (RMP=−66.1±3.30 mV) of 18 MDH neurons tested (RMP=−63.1±2.04 mV), with no response induced in nine neurons (RMP=−63.2±2.56 mV). Muscimol induced membrane depolarization in 4 (RMP=−59.2±6.04 mV) of 17 neurons tested in the IL-1β-treated group. Fig. 5C and 5D show comparisons of GABA and muscimol-induced responses between the control and IL-1β-treated groups, respectively. Interestingly, 18% (3/17) and 22% (4/18) of neurons tested showed GABA- and muscimol-induced membrane depolarization in the IL-1β-treated rats, respectively. However, none of the neurons tested in the control group were depolarized in response to GABA or muscimol. Collectively in the IL-1β-injected rats, GABA- and muscimol-induced membrane depolarization was observed, but there were no significant differences in the amount of depolarization between control and IL-1β-injected rats (χ2-test; GABA, p>0.05, Fig. 5C; muscimol, p>0.05, Fig. 5D).
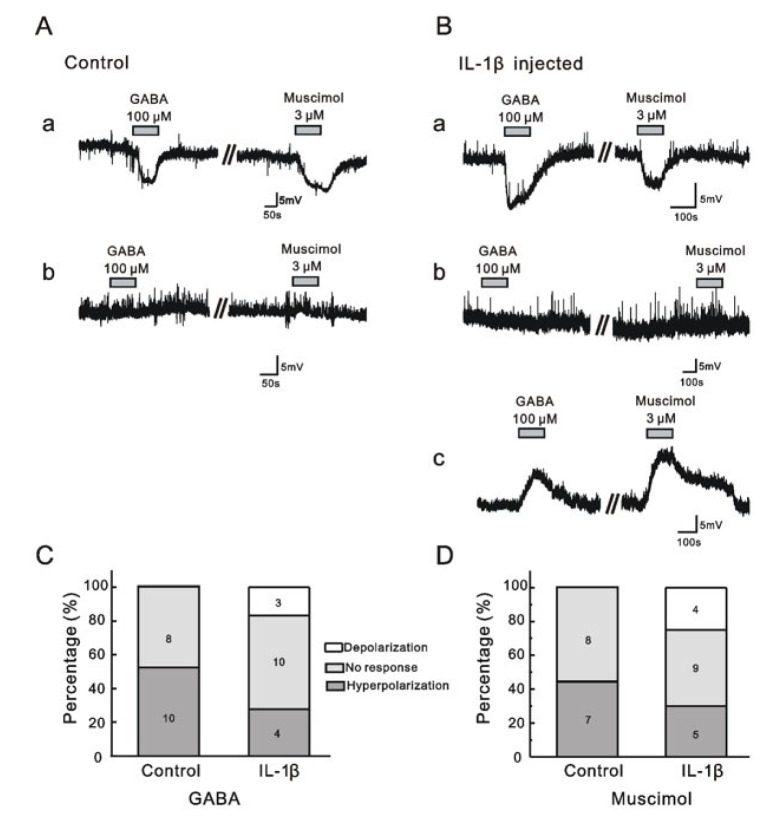 | Fig. 5GABA and muscimol-induced responses in MDH neurons in control and IL-1β-injected rats.(A) Representative traces showing membrane hyperpolarization (a) and no response (b) following GABA and muscimol addition to the membranes of neurons from control rats. (B) Representative traces showing membrane hyperpolarization (a), no response (b), and membrane depolarization (c) following GABA and muscimol addition to neuronal membranes of IL-1β-injected rats. (C and D) Bar graphs showing the proportions of membrane hyperpolarization, no response, and membrane depolarization following GABA (C) and muscimol (D) treatment in control and IL-1β-injected rats.
|
Go to :
DISCUSSION
Here we demonstrate that intracisternal administration of bicuculline, a GABAA receptor antagonist, leads to a paradoxical anti-allodynic effect in IL-1β-treated rats, whereas it leads to mechanical allodynia in naïve rats. Pretreatment with RTX, which depletes TRPV1 in primary afferent nerve fibers, did not affect the paradoxical anti-allodynic effects of bicuculline. Intracisternal injection of bumetanide, an NKCC1 inhibitor, reversed IL-1β-induced mechanical allodynia. In addition, we observed GABA- and muscimol-induced membrane depolarization in IL-1β-treated rats. These results suggest that some large myelinated Aβ fibers may gain access to the nociceptive system and result in pain sensation via GABAA receptors under inflammatory pain conditions.
It is well known that inhibition of central inhibitory neurotransmitters produces significant nociception. Several studies have demonstrated that acute antagonism of spinal GABAA receptors following bicuculline injection produces touch-evoked allodynia [2829]. The present study demonstrated that intracisternal administration of bicuculline produces mechanical allodynia and thermal hyperalgesia in the orofacial area of naïve rats. Although intracisternal administration of bicuculline produced mechanical allodynia and thermal hyperalgesia, the effective dose of bicuculline for mechanical allodynia was lower than that for thermal hyperalgesia. In contrast to what was observed in naïve rats, the present study observed that intracisternal administration of 50 ng of bicuculline produces anti-allodynic effects in IL-1β-treated rats without affecting IL-1β-induced thermal hyperalgesia. These results suggest that blockade of GABAA receptors leads to paradoxical anti-allodynic effects under inflammatory pain conditions. This conclusion is supported by previous studies, indicating that peripheral inflammation facilitates Aβ fiber-mediated synaptic input to MDH neurons of the adult rat spinal cord [30]. Moreover, inflammation of one hind paw in rats induced by carrageenan or complete Freund's adjuvant (CFA) leads to increases in GABA-immunoreactive cells [3132] and increased GABA release [31] in the spinal cord. Taken together, these results and those of the present study imply that central GABAA receptors participate in mediating mechanical allodynia produced by large myelinated Aβ fibers following inflammation.
Large myelinated primary afferent nerve fibers do not induce hyperalgesia or allodynia under normal conditions. Under inflammatory conditions, however, some large myelinated primary afferent nerve fibers might produce mechanical allodynia. A recent study reported that mechanical allodynia is mediated by large-diameter primary afferent nerve fibers whereas thermal hyperalgesia is mediated by small-diameter primary afferent nerve fibers under inflammatory conditions after subcutaneous injection of IL-1β [4]. Here we demonstrate that RTX treatment does not lead to alterations in bicuculline-induced paradoxical anti-allodynic effects in IL-1β-treated rats. These results suggest that small-diameter primary afferent nerve, which expressing TRPV1, do not mediate bicuculline-induced paradoxical anti-allodynic effects during inflammation. Thus, bicuculline-induced paradoxical anti-allodynic effects seem to be mediated by large-diameter primary afferent nerve fibers.
The present results imply that central GABAA receptormediated anti-nociception is produced by inhibitory action under normal conditions, while central GABAA receptor-mediated pro-nociception is produced by excitatory action under inflammatory conditions. These results are supported by previous data from electrophysiological studies. Removal of GABAergic inhibition facilitates polysynaptic A fiber-mediated excitatory transmission to the superficial spinal dorsal horn [33]. An important mechanism underlying this switch action from GABAergic inhibition to excitation may involve modifications in trans-membrane chloride ion concentrations [6]. The potassium chloride cotransporter (KCC) normally decreases the intracellular levels of potassium and chloride ions, whereas NKCC increases the intracellular levels of potassium and chloride ions. Both NKCCs and KCCs are chloride ion transporters in neurons and modulate the intracellular chloride ion concentrations.
Generally, bumetanide has an approximately 500-fold greater affinity for NKCC1 than for KCC2 [3435]. Our data indicate that intracisternal administration of a high dose of bumetanide (200 µg), but not low doses of bumetanide, decreases air-puff thresholds in naïve rats. This suggests that bumetanide also binds KCC2 at high doses. Therefore, low doses of bumetanide (50 and 100 µg) were used in the next experiment. Low doses of bumetanide reversed the decreased air-puff thresholds produced by IL-1β injection. These results suggest that NKCC1 is involved in touch-evoked allodynia under inflammatory conditions. The participation of NKCC1 in the switch from anti-nociception to pro-nociception is supported by several previous studies. In rats with arthritis, up-regulated expression of NKCC1 mRNA was observed in the superficial, but not deep spinal dorsal horn [36]. Moreover, an intracolonic capsaicin injection was shown to induce NKCC1 membrane delivery and phosphorylation in the dorsal spinal cord [37]. Therefore, up-regulated NKCC1 activity under inflammatory conditions leads to changes in the anion gradient in postsynaptic neurons, which can convert lead to the conversion of GABA-induced hyperpolarization to depolarization [36]. Previous studies also provide behavioral evidence for the involvement of NKCC1 in the conversion from GABA-induced hyperpolarization to depolarization. Intrathecal administration of bumetanide attenuated abdominal allodynia produced by intracolonic capsaicin injection [38] and phase II nociceptive behavioral responses in the formalin model [39]. These results, taken together with the present data, suggest that NKCC1 is involved in the altered spinal processing that leads to Aβ fiber-mediated touch-evoked allodynia under inflammatory pain conditions. Although there is evidence for increased NKCC1 expression [36], down-regulation of KCC2 in the spinal cord also contributes to neuropathic pain after peripheral nerve injury [4041] as well as inflammatory pain [4243]. Therefore, in order to fully elucidate the participation of KCC2 in Aβ fiber-mediated touch-evoked allodynia under inflammatory pain conditions, future studies will be required.
The present study provides evidence of a switch from GABA-mediated hyperpolarization to depolarization in gramicidin perforated current clamp mode in the MDH. No neurons with membrane depolarization were observed in control rats after GABA or muscimol application. However, application of GABA or muscimol produced membrane depolarization in 18% and 22% of neurons, respectively, in the IL-1β-treated rats, although there were no significant differences between the two groups. These results are in line with those of a previous study demonstrating that high expression of NKCC1 produces GABA-induced depolarization in newly generated granule cells in the dentate gyrus of adult mice and that GABA-induced depolarization is reversed by shRNA-mediated knockdown of NKCC1 expression [44]. These results, taken together with our recording data, suggest that GABA-mediated depolarization participates in the phenotypic switching of Aβ fibers, which provide an excitatory link from low-threshold inputs to nociceptive neurons in the trigeminal system under pathological pain conditions.
In summary, intracisternal administration of bicuculline produced a paradoxical anti-allodynic effect under inflammatory pain conditions. Intracisternal injection of bumetanide reversed the IL-1β-induced mechanical allodynia. Application of GABA or muscimol produced membrane depolarization in IL-1β-treated rats. These results suggest that NKCC1 in trigeminal neurons participates in a direct excitatory link between low-threshold inputs and nociceptive neurons by converting GABAA-induced hyperpolarization to depolarization under inflammatory pain conditions. Thus, some large myelinated Aβ fibers can gain access to the nociceptive system and elicit pain sensation under inflammatory conditions. Our present data suggest that modulation of GABAA receptor seems to be a potential new therapeutic target for inflammatory pain control.
Go to :
 XML Download
XML Download