

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Inflammation is a complex biological process that protects our body from invasion by various pathogens, such as bacteria, viruses, fungi, and other microorganisms. Inflammation is characterized by several hallmarks, including pain, redness, swelling, heat, and loss of function [1]. While inflammation is an innate immune response and a defense mechanism to protect our bodies against various dangers, chronic inflammation or repeated and prolonged inflammation has been regarded as a leading cause of various human diseases, including inflammatory/autoimmune diseases, such as rheumatoid arthritis, atherosclerosis, lupus erythematosus, and sepsis; metabolic diseases; and even cancers [23]. Several types of innate immune cells are actively involved in the inflammatory process; among these, macrophages are some of the most critical factors in inducing an inflammatory response.
Macrophages are the first defense cells to initiate an inflammatory response by detecting pathogens. Macrophages recognize conserved pathogen structures and molecules, known as pathogen-associated molecular patterns (PAMPs), such as cell wall components and nucleic acids, which are essential for pathogen survival and virulence [45]. This recognition is accomplished by molecular interaction between PAMPs and conserved pattern recognition receptors (PRRs) in macrophages, such as toll-like receptors (TLRs), RIG-I-like receptors (RLRs), and sensor proteins [678]. Through PAMP recognition by PRRs, especially TLRs, macrophages immediately induce inflammatory responses by releasing type I interferons (IFNs), such as IFN-α and -β, and proinflammatory cytokines, such as tumor necrosis factor (TNF)-α, interleukin (IL)-1b, and IL-6 [9101112]. Pro-inflammatory cytokines are thought to remove pathogens in an indirect manner, e.g., by recruiting more immune cells to inflamed lesions and killing pathogen-infected cells. On the other hand, type I IFNs remove pathogens in a more direct manner than do pro-inflammatory cytokines, e.g., by preventing pathogen replication and proliferation to reduce pathogen numbers. The molecular mechanism of the macrophage-mediated inflammatory response has been extensively studied. PAMPs binding to macrophage PRRs initiate rapid signal transduction cascades activating a variety of intracellular signaling molecules, such as Src; spleen tyrosine kinase (Syk); and mitogen-activated protein kinases (MAPKs), such as extracellular signal-regulated kinases (ERKs), c-Jun N-terminal kinase (JNK), and p38. These signaling molecules activate inflammatory transcription factors, such as nuclear factor-kappa-B (NF-κB) and activator protein (AP)-1 [1314151617]. Therefore, the intracellular signaling molecules in macrophages that are activated during inflammatory responses could be pharmaceutical targets to prevent and treat inflammatory and infectious diseases. Many studies have concentrated on identifying and testing novel anti-inflammatory and anti-infectious agents targeting signaling molecules in macrophages.
Hydroquinone (benzene-1,4-diol; HQ; Fig. 1) is a benzenederived metabolite that is a major component of industrial chemicals; cigarette smoke; and some foods, such as coffee, tea, and beer [1819]. Because cigarette smoking induces allergenic and inflammatory reactions, possibly due to the presence of benzene metabolites, which are strong haptens [20], previous studies have demonstrated HQ to be an immunotoxicological substance in allergenic and inflammatory diseases [21]. A number of recent studies have reported anti-inflammatory activities of HQ, including suppression of the expression of pro-inflammatory cytokines, such as TNF-α, IL-1β, and IL-6, by targeting AKT, which is a critical signaling molecule in the NF-kB pathway in lipopolysaccharide (LPS)-activated macrophages [2223242526]. In addition, HQ suppressed the production and release of inflammatory mediators, such as nitric oxide (NO) and prostaglandin E2 (PGE2), and ameliorated hepatitis symptoms in an LPS/D-galactosamine-induced hepatitis mouse model [27]. These results strongly suggest that HQ and possibly benzene-derived metabolites are not simple toxic agents, but potential immunosuppressants with anti-inflammatory activities in macrophage-mediated inflammation. Despite these previous studies, new anti-inflammatory roles of HQ and the molecular mechanisms underlying macrophage-mediated inflammatory responses are still poorly understood.
In this study, we investigated a new role of HQ during inflammatory responses and the molecular mechanisms by which HQ modulates inflammatory responses in macrophages using a well-established, in vitro-activated macrophage platform, LPS-stimulated RAW264.7 cells. The results of this study provide strong evidence of a previously unknown role of HQ in suppressing the expression of a type I IFN, IFN-β, a critical cytokine in preventing pathogen proliferation, in macrophages by targeting AKT kinases in the interferon regulatory factor (IRF)-3 signaling pathway.
Go to : 
METHODS
Materials
HQ (Fig. 1), lipopolysaccharide (LPS, E. coli 0111:B4), 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT), and polyethylenimine (PEI) were purchased from Sigma Chemical Co. (St. Louis, MO, USA). RAW264.7 and HEK293 cells were purchased from ATCC (Rockville, MD, USA). Roswell Park Memorial Institute (RPMI) 1640, Dulbecco's modified Eagle's medium (DMEM), fetal bovine serum (FBS), phosphate-buffered saline (PBS), streptomycin, penicillin, and L-glutamine were purchased from Gibco (Grand Island, NY, USA). TRI reagent® was purchased from Molecular Research Center Inc. (Cincinnati, OH, USA). MuLV reverse transcriptase was purchased from Thermo-Fisher Scientific (Waltham, MA, USA). Primers used for semiquantitative polymerase chain reaction (PCR) were synthesized, and PCR premix was purchased from Bioneer Inc. (Daejeon, Korea). Antibodies specific for phosphorylated and total forms of IRF-3, TBK-1, AKT, hemagglutinin (HA) tag, and β-actin were purchased from Cell Signaling Technology (Beverly, MA, USA). Enhanced chemiluminescence system was purchased from AbFrontier (Seoul, Korea). AKT gene constructs (AKT1, AKT2, AKT3, and AKT KD) used in this study were purchased from Addgene (Cambridge, MA, USA). TRIF and TKB1 constructs were used as previously reported [28].
Cell culture
RAW264.7 and HEK293 cells were maintained in RPMI 1640 medium and DMEM, respectively, containing 10% heat-inactivated FBS, 100 mg/ml streptomycin, 100 U/ml penicillin, and 2 mM L-glutamine at 37℃ in a 5% CO2 humidified incubator. The cells were subcultured with new cell culture media three times a week to maintain the cells.
Semi-quantitative PCR
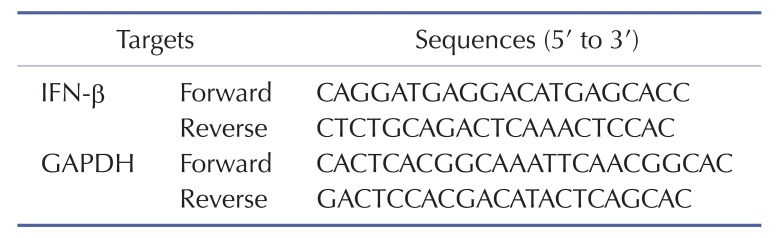
To determine the level of IFN-β mRNA expression, RAW264.7 (1×106 cells/ml) cells were pretreated with escalating doses of HQ (0, 25, 50, and 75 µM) for 30 min and then incubated with LPS (1 µg/ml) for 6 h. Total RNA was extracted with TRI reagent® according to the manufacturer's instructions. cDNA was immediately synthesized from 1 mg of total RNA by MuLV reverse transcriptase according to the manufacturer's instructions and used for semi-quantitative PCR as previously described [329]. The primer sequences for the semi-quantitative PCR are listed in Table 1.
IRF-3-luciferase reporter gene assay
HEK293 cells (1×106 cells/ml) were co-transfected with IFN-β-Luc and β-gal along with TIR-domain-containing adapter-inducing interferon-β (TRIF), TANK-binding kinase 1 (TBK1), or an AKT construct for 24 h using PEI in 24 well plates and treated with escalating doses of HQ (0, 25, 50, and 75 µM) for another 24 h. The cells were next lysed by freezing and thawing, and the luciferase activity in the lysate was measured with a luciferase assay as reported previously [33031]. All luciferase activities were normalized to b-galactosidase activities.
Cell viability assay
The cytotoxic effect of HQ on RAW264.7 cells (1×106 cells/ml) was determined by treating cells with escalating doses of HQ (0, 5, 10, 25, 50, and 75 µM) for 24 h and performing an MTT assay as previously reported [3233]. Briefly, cell culture medium (100 ml) was mixed with MTT solution (10 µl; 10 mg/ml in PBS, pH 7.4) and incubated for 4 h at 37℃. A 15% sodium dodecyl sulfate solution was directly added to the mixture to stop the reaction, and the mixture was further incubated at 37℃ for 24 h. The OD at 570 nm was determined using a Spectramax 250 microplate reader.
Preparation of total cell lysates and Western blot analysis
Total lysates of RAW264.7 cells were prepared as previously described [3134]. Briefly, RAW264.7 cells were pretreated with the indicated doses of HQ, incubated with LPS (1 µg/ml) for the indicated time, and washed three times with PBS. The cells were lysed by a sonication on ice for 1 min in cold cell lysis buffer (20 mM Tris–HCl, pH 7.4, 2 mM EDTA, 2 mM ethyleneglycotetraacetic acid, 50 mM β-glycerophosphate, 1 mM sodium orthovanadate, 1 mM dithiothreitol, 1% Triton X-100, 10% glycerol, 10 µg/ml aprotinin, 10 µg/ml pepstatin, 1 mM benzimide, and 2 mM PMSF). Clear total lysates were collected by centrifugation at 16,000 × g for 10 min at 4℃, discarding the pellet, and storing immediately at −80℃ until use. For Western blot analysis, total cell lysates were subjected to SDS-polyacrylamide electrophoresis and transferred to polyvinylidene fluoride membrane at 4℃ and 250 mA for 1 h. Total and phosphorylated forms of target proteins IRF-3, TBK1, AKTs, and β-actin were detected by antibodies specific for each target and visualized with an enhanced chemiluminescence system according to the manufacturer's instructions as reported previously [35].
Statistical analysis
All data in this study are presented as the mean and standard deviation of at least three independent experiments. To determine statistical significance among experimental groups, all results were analyzed by ANOVA/Scheffe's post hoc test and Kruskal-Wallis/Mann-Whitney test. p<0.05 was considered statistically significant, and all statistical analyses were performed with SPSS.
Go to :
RESULTS AND DISCUSSION
Previous studies have reported that HQ has various immunomodulatory activities. HQ has been shown to ameliorate the symptoms of allergic diseases by regulating immune responses. HQ significantly reduced IL-12 production in LPS-stimulated macrophages, which suppress immune responses mediated by type-2 helper T-cells associated with the development of allergic disease by suppressing NF-κB activity [21]. In addition, HQ reduced the production of some cytokines and cytotoxic molecules, such as IL-2, IL-1β, and NO, in macrophages [2226]. HQ was also reported to induce IL-4 production from CD4+ T-cells and increase serum immunoglobulin E level in antigen-primed mice [36]. Recent studies have demonstrated that HQ is also actively involved in modulating inflammatory responses in macrophages. HQ significantly decreased the production of pro-inflammatory cytokines, including TNF-α, IL-1β, and IL-6, as well as inflammatory molecules, such as NO, in LPS-activated macrophages by suppressing the NF-kB signaling pathway [22242537]. Although these studies suggest that HQ modulates a wide range of immune responses in immune cells, the role of HQ in immune responses, especially macrophage-mediated inflammatory responses, is poorly understood. Therefore, we investigated the role of HQ in macrophage-induced inflammatory responses and the underlying molecular mechanism using LPS-stimulated RAW264.7 cells.
Upon recognizing invading pathogens, macrophages immediately initiate inflammatory responses in a cooperative manner by producing pro-inflammatory cytokines, such as TNF-α, IL-1β, and IL-6, and anti-pathogen cytokines, such as type I IFNs. Pro-inflammatory cytokines help remove invading pathogens indirectly by recruiting circulating immune cells to the inflamed lesions through vasodilation and killing pathogen-infected cells. Type I IFNs directly remove invading pathogens by suppressing transcription and translation of pathogen components to inhibit pathogen replication and proliferation, resulting in pathogen removal. Because previous studies have successfully demonstrated that HQ significantly reduced the production of pro-inflammatory cytokines and molecules [2225], the effect of HQ on IFN-β production was examined in the present study. HQ suppressed IFN-β mRNA expression in a dose-dependent manner in LPS-stimulated RAW264.7 cells (Fig. 2A) but did not have any cytotoxic effects at low (5 µM) to high (75 µM) doses (Fig. 2B). These results indicate that decreased IFN-b production due to HQ is not a result of cytotoxicity, but is due to HQ-specific inhibition of IFN-β transcription. Since HQ decreased IFN-β transcription in LPS-stimulated RAW264.7 cells, we examined the effects of HQ on the transcriptional activity of IRF-3, the key transcription factor inducing IFN-β expression, using a luciferase reporter gene. TRIF and TBK1 have been reported to induce IFN-β mRNA expression by activating IRF-3 [1538], and TRIF and TBK1 induced IRF-3-mediated luciferase activity in HEK293 cells. HQ at all doses significantly suppressed TRIF-induced, IRF-3-mediated luciferase activity (Fig. 2C; left panel), but it did not suppress TBK1-induced, IRF-3-mediated luciferase activity (Fig. 2C; right panel). Although HQ down-regulated IFN-β mRNA expression in a dose-dependent manner (Fig. 2A), suppression of IRF-3-mediated luciferase activity by HQ was not dose dependent; all HQ doses statistically significantly suppressed IRF-3-mediated luciferase activity. Why the effects of HQ on IFN-β mRNA expression and IRF-3-mediated luciferase activity are not consistent remains unclear, so further study is required.
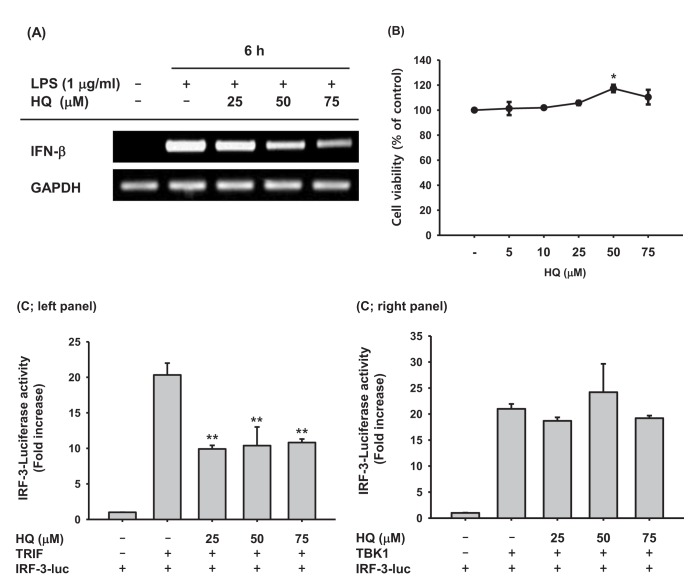 | Fig. 2Effect of HQ on the mRNA expression of IFN-β(A) RAW264.7 cells (1×106 cells/ml) pretreated with the indicated doses of HQ for 30 min were incubated with LPS (1 µg/ml) for 6 h. Expression levels of IFN-β and GAPDH mRNA were determined by semi-quantitative PCR. (B) RAW264.7 cells (1×106 cells/ml) were treated with the indicated doses of HQ for 24 h, and cell viability was determined by MTT assay. (C) HEK293 cells (1×106 cells/ml) cotransfected with IRF-3-Luc (1.0 µg/ml), β-gal (0.1 µg/ml), and either (Left panel) TRIF (1.0 µg/ml) or TBK1 (1.0 µg/ml) for 24 h were treated with the indicated doses of HQ for another 24 h. Luciferase activity was measured with a luminometer. All luciferase activities were normalized to β-galactosidase activity. *p<0.05, **p<0.01 compared to the control.
|
Since HQ effectively suppressed IRF-3 activity induced by TRIF and TBK1 (Fig. 2C), we next investigated the effect of HQ on IRF-3 activity in macrophages by Western blot analysis. The effect of HQ on IRF-3 activity was examined in LPS-stimulated RAW264.7 cells, and HQ dramatically reduced IRF-3 phosphorylation at 15 and 30 min after LPS treatment in RAW264.7 cells (Fig. 3A). To confirm this result, the effect of HQ on IRF-3 activity was examined in TRIF-transfected RAW264.7 cells since HQ significantly suppressed IRF-3-mediated luciferase activity induced by TRIF more than that induced by TBK1 (Fig. 2C). As expected, HQ at all doses effectively reduced IRF-3 phosphorylation in TRIF-transfected RAW264.7 cells (Fig. 3B). These results strongly suggest that HQ suppresses IRF-3 activation by inhibiting its phosphorylation and consequently decreases IRF-3 activity, resulting in down-regulated IFN-β mRNA expression in LPS-stimulated RAW264.7 cells.
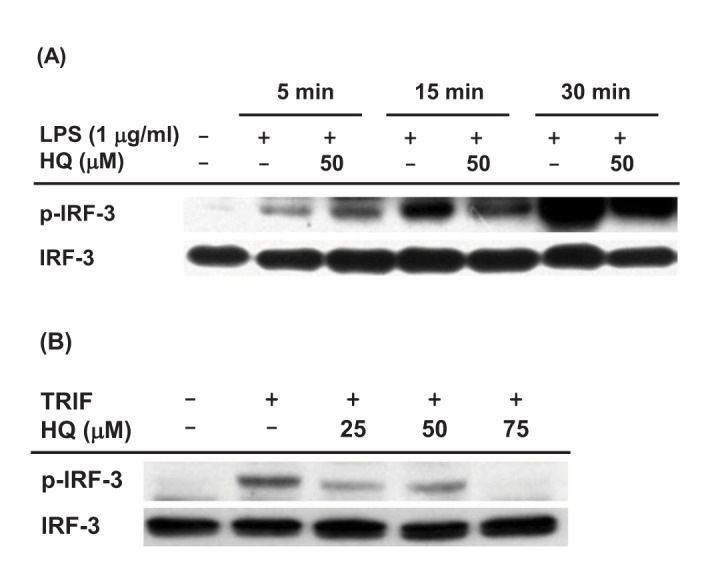 | Fig. 3Effect of HQ on the activation of IRF-3(A) RAW264.7 cells (1×106 cells/ml) pretreated with either HQ (50 µM) or vehicle for 30 min were incubated with LPS (1.0 µg/ml) for the indicated time. Phosphorylated and total levels of IRF-3 in total cell lysates were determined by Western blot analysis. (B) RAW264.7 cells (1×106 cells/ml) transfected with TRIF (1.0 µg/ml) for 24 h were treated with the indicated doses of HQ for 24 h, and phosphorylated and total levels of IRF-3 in total cell lysates were determined by Western blot analysis.
|
A number of previous studies have established that many intracellular molecules in the IRF-3 signaling pathway, such as AKT, TRIF, TNF receptor associated factor 3 (TRAF3), TNAK, TBK1, and inhibitor of nuclear factor kappa-B kinase ε (IKKε), are activated in IRF-3 signaling [1415]. Since HQ suppressed IRF-3 activation in LPS-stimulated RAW264.7 cells, we next analyzed the upstream signaling molecules targeted by HQ. To examine which intracellular molecules in the IRF-3 signaling pathway are targeted by HQ, the phosphorylation of critical upstream molecules in the IRF-3 signaling pathway, such as TBK1 and AKTs, were determined in the LPS-stimulated RAW264.7 cells treated with HQ. HQ dramatically reduced AKT phosphorylation from 15 to 30 min. On the other hand, HQ did not suppress TBK1 phosphorylation in LPS-stimulated RAW264.7 cells (Fig. 4A). This result is consistent with previous findings that HQ did not suppress the IRF-3-mediated luciferase activity in TBK1-transfected cells (Fig. 2C right panel) and suggests that HQ suppresses IRF-3 activity by targeting AKT rather than TBK1. To confirm that HQ suppressed AKT phosphorylation (Fig. 4A), RAW264.7 cells transfected with AKT were treated with HQ, and IRF-3 phosphorylated was examined by Western blot. As expected, IRF-3 phosphorylation induced by AKT transfection was dramatically decreased by HQ (Fig. 4B). To further confirm that AKT is a target for HQ-mediated suppression of IRF-3 activation in LPS-stimulated RAW264.7 cells, RAW264.7 cells were transfected with either wild-type AKT or mutant AKT with cysteine 310 substituted for alanine (AKT C310A). HQ directly targets and suppresses AKT phosphorylation at both threonine 308 and serine 473 [24]. IRF-3 phosphorylation in these cells was examined by Western blot analysis. Wild-type AKT dramatically induced IRF-3 phosphorylation in RAW264.7 cells, whereas AKT C310A poorly increased IRF-3 phosphorylation (Fig. 4C). These results indicate that HQ suppresses IRF-3 activation by targeting AKT at cysteine 310 in LPS-stimulated RAW264.7 cells. To date, three different AKT isoforms (AKT1, AKT2, and AKT3) have been identified in mice and humans, but their roles in modulating immune responses have not yet been elucidates. To determine which AKT is the target for HQ-mediated inhibition of IRF-3 activation in LPS-stimulated macrophages, RAW264.7 cells were transfected with each AKT isoform in the absence or presence of HQ, and IRF-3 phosphorylation was examined by Western blot. HQ inhibited phosphorylation of all three AKTs, resulting in suppressed IRF-3 phosphorylation in all RAW264.7 cells transfected with AKT isoforms (Fig. 4D). As previously reported, HQ suppresses AKT1 activation by targeting cysteine 310, abolishing phosphorylation at threonine 308 and serine 473 [24]. This mechanism was confirmed for HQ-mediated suppression of IRF-3 activation in LPS-stimulated macrophages (Fig. 4C). Cysteine 310 is evolutionarily well conserved among AKT isoforms. AKT2 and AKT3 have cysteine residues at 311 and 307, respectively, suggesting that HQ targets cysteine 311 of AKT2 and cysteine 307 of AKT3 to suppress AKT phosphorylation and IRF-3 activity in LPS-stimulated macrophages. Although this mechanism is reasonable, it needs to be confirmed in further investigation. Since HQ suppressed phosphorylation of all three AKT isoforms and reduced IRF-3 phosphorylation in LPS-stimulated RAW264.7 cells, we further investigated the effect of HQ on the suppression of IRF-3 activation with different AKT isoforms using a luciferase reporter gene. IRF-3-mediated luciferase activities were determined in HEK293 cells transfected with each AKT isoform in the absence or presence of HQ, and HQ significantly suppressed the IRF-3-mediated luciferase activities induced by each AKT (Fig. 4E). These results strongly indicate that HQ suppresses IRF-3 activation, which is critical for expression of IFN-b mRNA, by targeting all three AKT isoforms in LPS-stimulated macrophages.
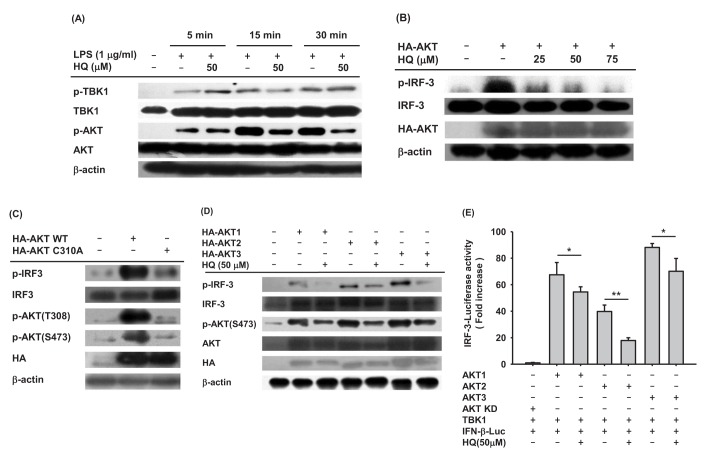 | Fig. 4Effect of HQ on AKT activation in IRF3 pathway(A) RAW264.7 cells (1×106 cells/ml) pretreated with either HQ (50 µM) or vehicle for 30 min were incubated with LPS (1.0 µg/ml) for the indicated time. Phosphorylated and total levels of TBK1 and AKT in total cell lysates were determined by Western blot analysis. (B) RAW264.7 cells (1×106 cells/ml) transfected with HA-AKT (1.0 µg/ml) for 24 h were treated with the indicated doses of HQ for another 24 h, and phosphorylated and total levels of IRF-3 and HA in total cell lysates were determined by Western blot analysis. (C) RAW264.7 cells (1×106 cells/ml) were transfected with either HA-AKT (1.0 µg/ml) or HA-AKT C310A (1.0 µg/ml) for 48 h. Phosphorylated and total levels of IRF-3, AKT, and HA in total cell lysates were determined by Western blot analysis. (D) RAW264.7 cells (1×106 cells/ml) transfected with either HA-AKT1 (1.0 µg/ml), HA-AKT2 (1.0 µg/ml), or HA-AKT3 (1.0 µg/ml) for 24 h were treated with either HQ (50 µM) or vehicle for another 24 h. Phosphorylated and total levels of IRF-3, AKT, and HA in the total cell lysates were determined by Western blot analysis. (E) HEK293 cells (1×106 cells/ml) cotransfected with TBK1 (1.0 µg/ml), IRF-3-Luc (1.0 µg/ml), β-gal (0.1 µg/ml), and HA-AKT1 (1.0 µg/ml), HA-AKT2 (1.0 µg/ml), HA-AKT3 (1.0 µg/ml), or HA-AKT KD (kinase domain deletion mutant) for 24 h were treated with either HQ (50 µM) or vehicle for another 24 h. Luciferase activity was measured with a luminometer. All luciferase reporter gene activities were normalized to β-galactosidase activity. *p<0.05, **p<0.01 compared to the control.
|
Go to :
CONCLUSION
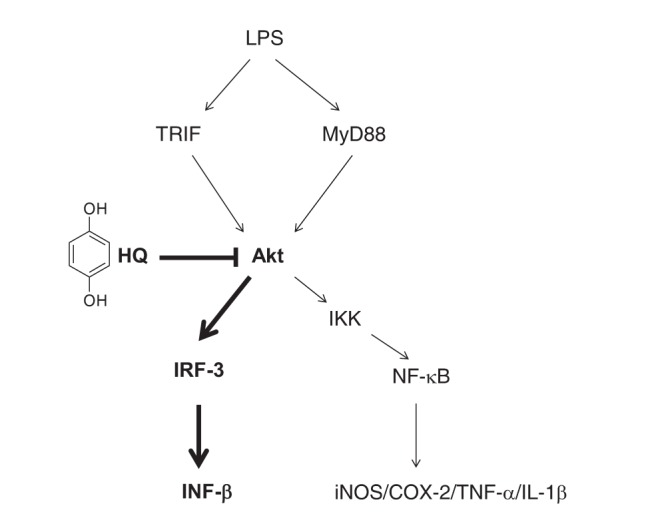
The present study aimed to explore the anti-inflammatory activity of HQ in activated macrophages and provided new evidence that HQ effectively suppresses expression of IFN-β mRNA, which plays a crucial role in inhibiting pathogens from replicating and proliferating in host cells. This inhibitory activity was accomplished by targeting AKTs, which are critical upstream kinases in the IRF-3 signaling pathway, suppressing IRF-3 activity, which is required for macrophages to produce type I IFNs, as described in Fig. 5. These results suggest that HQ could be a novel agent to prevent and treat inflammatory and infectious diseases by preventing the production of type I IFNs during macrophage-mediated inflammatory responses.
Go to :
 XML Download
XML Download