

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Dipeptidyl peptidase 4 (DPP-4) is widely expressed on the surface of many cell types and is involved in the dipeptidyl enzymatic metabolism of incretin hormones [1]. The dipeptidyl peptidase enzyme activity is blocked by DPP-4 inhibitors for the treatment of type 2 diabetes mellitus (T2DM) [2]. However, the factors and mechanisms that control DPP-4 activity are very complex and need to be clarified further; hence, further studies are required even after ten years of widespread use of DPP-4 inhibitors in diabetes clinics [3]. Further, as it is associated with insulin resistance and diabetes, DPP-4 expression is increased in adipose tissue and its stromal vascular dendritic cells/macrophages [4] and peripheral blood T cells [5]. Some studies have shown that glucose level affects DPP-4 activity and expression per se [6]. We have also reported that T cell DPP-4 expression, serum soluble DPP-4 and DPP-4 activity were increased in patients with T2DM, and the increase was related to glycemic status [5]. Also, DPP-4 can be regulated in a cell- or tissue-specific manner. In Caco2 cells, high glucose concentrations suppress DPP-4 gene expression, resulting in decreased DPP-4 enzymatic activity [7]. In HepG2 cells, DPP-4 activity and gene expression were found to be increased by high dose of sodium butyrate (SB) [8].
Recently, butyric acid and its solute form SB have been gaining increasing attention due to the metabolic actions of short-chain fatty acids (SCFAs), which are naturally produced in the distal small intestine and colon through fermentation of resistant starch, dietary fiber, and other low-digestible polysaccharides [9]. SB and other SCFAs, suppress histone deacetylases (HDACs), thereby regulating cell growth, differentiation and apoptosis in multiple cells [10]. In dietary obese C57BL/6J mice, SB was shown to suppress weight gain independent of food intake suppression. Also, except for the intestinal lumen, most of SCFAs in tissue and blood exist in their submolar concentrations [11], which is in contrast to the high molar concentration of SB that was used in in vitro experiments. Many studies have suggested that butyrate has pleiotropic effects through the modification of chromatin plasticity, histone acetylation, histone phosphorylation and/or phosphorylation of high-mobility group proteins [8]. However, some studies have shown that SB has dual effects on cell growth, cell cycle distribution, and gene expression in hepatocellular carcinoma cells depending on the concentration [12] and cell-specific actions depending on the cell type [13], which warrant further studies regarding the previously reported effect of SB on DPP-4. To understand clinical implications of the pleiotropic actions of butyrate, it seems necessary to address the context-dependent actions of butyrate. In this study, we evaluated the effect of SB on regulation of DPP-4 activity and expression in cells and high fat diet (HFD)-induced obese mice to better characterize the metabolic actions of SB.
METHODS
Materials
RPMI 1640, fetal bovine serum (FBS) and other cell culture reagents were purchased from Gibco BRL Co. (Grand Island, NY, USA). All primary and secondary antibodies, unless otherwise stated, were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Anti-acetyl-H3 and anti-acetyl-H4 antibodies were purchased from Millipore (Bedford, MA, USA). The enzyme-linked immunosorbent assay (ELISA) kits for interleukin (IL)-1β, IL-6, and tumor necrosis factor-α (TNF-α) were purchased from R&D Systems (Minneapolis, MN, USA). Histone extraction kit was obtained from Abcam (Cambridge, UK). Other chemicals, unless stated otherwise, were obtained from Sigma Chemical Co. (St. Louis, MO, USA).
Cell culture
HepG2 and mesangial cells were obtained from the American Type Culture Collection (ATCC, Manassas, VA, USA). HepG2 cells were maintained in RPMI 1640 supplemented with 10% heat-inactivated fetal bovine serum (FBS), penicillin G (100 units/mL), streptomycin (100 mg/mL), and L-glutamine (2 mmol/L), at 37℃ in a humidified atmosphere containing 5% CO2. Mouse mesangial cells (ATCC, Manassas, VA, USA) were maintained in Dulbecco's Modified Eagle Medium: Nutrient Mixture F-12 (DMEM/F12) supplemented with 10% heat-inactivated fetal bovine serum (FBS), with other culture conditions being similar to HepG2 cells. Most experiments were performed in mesangial cells after 3 days of incubation with SB, unless stated otherwise.
Animal experiments
The animal procedures were conducted in accordance with the 1985 (revised 1996) Guidelines for the Care and Use of Laboratory Animals of the U.S. National Institutes of Health and were approved by the Institutional Animal Care and Utilization Committee for Medical Science of Wonkwang University (Approval NO. WKU15-44). C57BL/6J mice (n≥9 in each group) were obtained from Orient Bio (Sungnam, Kyungkido, South Korea). High-fat diet (HFD; 60% of total calories) was purchased from Research Diets, Inc (New Brunswick, NJ, USA, Cat. D12492). All animals were bred and housed in standard cages in a climate-controlled room at an ambient temperature of 23±2℃. C57BL/6J mice were fed normal chow diet (NCD) or HFD for 10 weeks. Then, HFD-induced obese mice were divided into two groups: HFD only or HFD+SB. In the “HFD+SB” group, mice were kept on HFD with SB incorporated at 5% wt/wt (n=9), while in the “HFD only” group (HFD, n=9) and the “control” group (Control, n=9), mice were kept on the same HFD or NCD, respectively. At 16 weeks thereafter, mice from the three experimental groups were sacrificed. Blood and tissues including liver and kidney were collected appropriately. To prepare plasma for the measurement of active GLP-1, we collected blood in ice-cooled ethylenediaminetetraacetic acid (EDTA)-plasma tubes which contained a DPP-4 inhibitor (10 µl per milliliter of blood) obtained from EMD Millipore Corporation (St. Charles, Missouri, USA) and we mixed the blood gently with the reagents. The tubes were maintained in an ice bath and centrifuged within one hour at 1000 x g for 10 minutes in a refrigerated centrifuge. Specimens were stored at −70 ℃ until assayed.
Western blot analysis
Cells or mouse tissues were washed with PBS and lysed using RIPA lysis buffer containing 25 mmol/L Tris-HCl buffer (pH 7.6), 150 mmol/L NaCl, 1% NP-40, 1% sodium deoxycholate, and 0.1% SDS. The protein concentration was determined using Bradford Assay Reagent (Bio-Rad, PA, USA). An equal amount of protein from each sample was resolved using sodium dodecyl sulfate-polyacrylamide gel electrophoresis and it was then electrophoretically transferred onto a Hybond-enhanced chemiluminescence nitrocellulose membrane (Bio-Rad, PA, USA). The membrane was blocked with 5% skim milk and incubated with primary antibodies at 4℃ overnight. The bands were visualized with enhanced chemiluminescence and quantified by densitometry. Representative blots from three independent experiments are presented in the figures. Nuclear and cytoplasmic extracts of cells were prepared using NE-PER nuclear and cytoplasmic extraction reagents (Pierce Biotechnology, Rockford, IL USA), respectively.
DPP-4 activity assay
DPP-4 activity was determined according to the cleavage rate of p-nitroaniline (pNA; Sigma) using the synthetic substrate H-glycyl-prolyl-p-nitroanilide (Gly-Pro-pNA; Sigma) [14]. Briefly, 100 µl of assay mixture was prepared containing 50 mM Tris (pH 8.0) and the protein sample from cells or mice tissues, or cell culture medium. The reaction was initiated by the addition of a 5 mM Gly-Pro-pNA substrate solution to a final concentration of 2.5 mM. After incubation for 10~30 min at 37℃, the absorbance of the sample in each well was measured at 405 nm using a plate reader. DPP-4 activity was expressed as the amount of cleaved pNA released per minute per ml (nmol/min/ml) for the culture medium and as the amount of pNA released per minute per weight (nmol/min/mg) for cells/tissues.
Real-time PCR
RNA was extracted from cells or tissues using an RNeasy Mini Kit (Qiagen). For real-time polymerase chain reaction (PCR), first-strand complementary DNA (cDNA) was synthesized from 1 µg total RNA using an Advantage RT-for-PCR Kit (Takara Korea Biomedical Inc., Seoul, Korea). Relative messenger RNA levels were determined by real-time PCR using a Brilliant II SYBR Green QPCR Master Mix Kit (Stratagene) and an Mx3000P thermal cycler (Stratagene). All cDNA levels were normalized to the level of glyceraldehyde 3-phosphate dehydrogenase (GAPDH). The primer sequences were designed using PrimerQuest (Integrated DNA Technologies, Cambridge, MA, USA).
Cytokine and monocyte chemotactic protein (MCP)-1 measurements
The serum levels of TNF-α, IL-1β, IL-6, and MCP-1 in each sample were determined using a commercially available kit from R&D Systems (Minneapolis, MN, USA). The assay was performed according to the manufacturer's instructions.
Histone extraction
Histone was extracted from cells using a commercially available Histone extraction kit from Abcam (Cambridge, UK). The assay was performed according to the manufacturer's instructions.
Statistical analysis
The data were expressed as the mean±standard error (S.E.) of at least three independent experiments or at least five independent samples and for at least six mice in each group. To compare three or more groups, one-way analysis of variance (ANOVA) followed by the Newman-Keuls post hoc test was used. Statistical analysis was performed using GraphPad Prism software, version 3.03 (GraphPad Software Inc., San Diego, CA, USA).
RESULTS
SB has dual effects on regulation of DPP-4 in HepG2 cells
To evaluate the effect of SCFAs on DPP-4 activity, we incubated HepG2 cells with SB, sodium acetate (SA), or sodium propionate (SP) for 3 days and measured DPP-4 activity in cell lysates. SB inhibited DPP-4 activity at the concentration range of 1 to 100 µM, whereas the DPP-4 activity was increased when the cells were treated with high concentration (1,000 µM) of SB, at which HDAC activity was reported to be inhibited significantly [12]. SP treatment of HepG2 cells showed similar but weaker effects on DPP-4 activity compared with SB. However, SA increased DPP-4 activity in HepG2 cells at 1 and 10 µM concentration. After we observed prominent and reciprocal effects of SB on DPP-4 activity in HepG2 cells, we focused on the effect of SB and evaluated the effects of SB on mRNA and protein expressions of DPP-4 and the acetylation of histone proteins in HepG2 cells. The expressions of both DPP-4 protein and mRNA were significantly increased in HepG2 cells treated with high concentration of SB, whereas both DPP-4 protein and mRNA expressions were down-regulated in HepG2 cells treated with a lower concentration of SB (100 µM) (Figs .1A–1E). We also investigated the expression of acetylated histone 3 (Acetyl-H3) and acetylated histone 4 (Acetyl-H4), as surrogate markers of HDAC inhibition, and we found the expression levels of Acetyl-H3 and Acetyl-H4 were consistently increased in HepG2 cells treated with high concentration of SB, without suppression at a lower concentration of SB (Fig. 1F). These results suggest that the suppression of DPP-4 activity and expression in HepG2 cells by relatively lower concentrations of SB is not related to the inhibitory action of SB on HDAC.
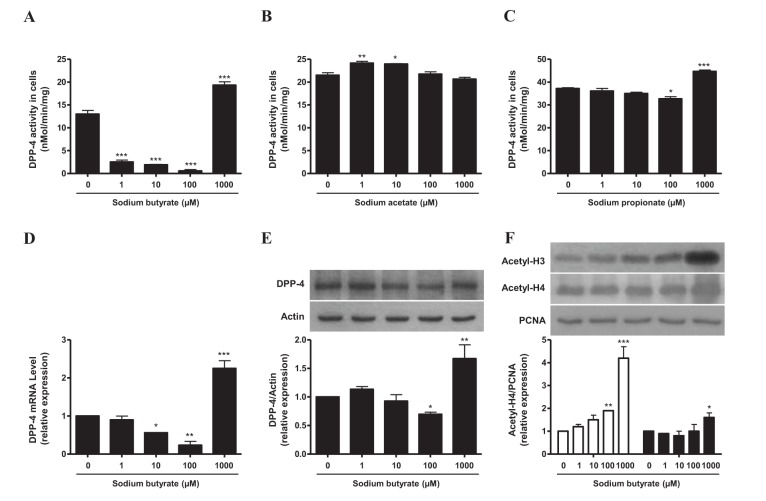
Fig. 1
The effects of SB on DPP-4 regulation (A~E) or acetyl-H3 and acetyl-H4 acetylation (F) in HepG2 cells.
HepG2 cells were treated with SB (0, 1, 10, 100, 1,000 µM) for 72 h. DPP-4 activity was determined as described in the “Methods” section. Histone was extracted using a histone extraction kit. Data represent means±S.E. *p<0.05, **p<0.01, and ***p<0.001 vs. untreated cells.
![]()
SB does not suppress DPP-4 activity in HepG2 cells under high glucose condition
Our previous study showed that serum DPP-4 activity and soluble DPP-4 level were increased in patients with T2DM [5]. These previous findings prompted us to evaluate whether the exposure of HepG2 cells to high glucose increases DPP-4 activity or not. DPP-4 activity and mRNA expression were markedly increased when the cells were incubated for 6 or more days under high glucose (30 mM) condition compared with that in HepG2 cells incubated in control media containing 11 mM glucose (Figs. 2A–2C). To explore if SB has a suppressive effect on the high glucose-induced increase in DPP-4 activity, we incubated HepG2 cells in high glucose media supplemented with SB. Interestingly, low concentration of SB did not suppress DPP-4 activity under high glucose condition. However, contrary to the findings in control media, SB at the low concentration range increased DPP-4 activity in HepG2 cells under high glucose conditions within 2~6 days (Figs. 2D–2F). The results suggest that SB, at its physiologically relevant concentration, has a context-dependent effect on DPP-4, at least in HepG2 cells.
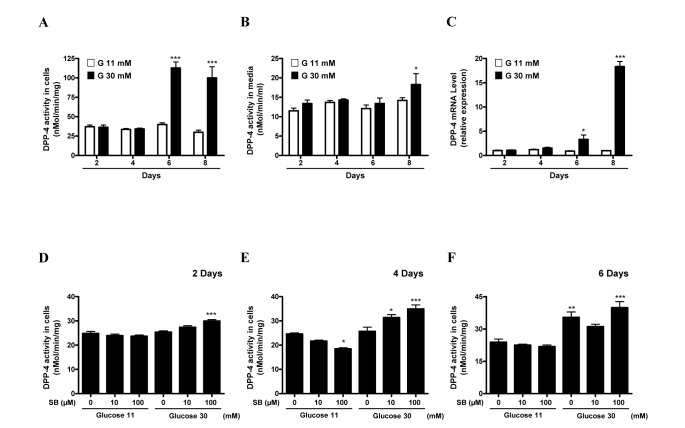
Fig. 2
The effects of SB on DPP-4 regulation in HepG2 cells and culture medium under co-stimulation with normal or high glucose condition (A~F).
HepG2 cells were treated with glucose (11 and 30 mM) for up to 8 days (A~C). HepG2 cells were treated with glucose (11 and 30 mM) for 2 days (D), 4 days (E), and 6 days (F). DPP-4 activity was determined as described in the “Methods” section. Data represent means±S.E. *p<0.05, **p<0.01 ***p<0.001 vs. glucose 11 mM treated cells.
Figure
![]()
SB suppresses DPP-4 activity and improves metabolic abnormalities and inflammatory cytokine profiles in HFD-induced obese mice
Before studying the in vivo effect of SB on HFD-induced obese mice, we checked the serum DPP-4 activity in C57BL/6J mice that were fed HFD for different periods from 0 to 24 weeks. Each set of mice (n=6 each) fed HFD for the indicated time period, as shown in Fig. 3A, was matched for age at sacrifice to exclude the effect of ageing on DPP-4 activity. Serum DPP-4 activity was increased significantly from 6 weeks of HFD in the experimental mice, and thereafter continuously across the period of HFD feeding (Fig. 3A). Then, to evaluate the metabolic effect of SB on DPP-4, and in obesity and insulin resistance, we administered SB to HFD-induced obese mice that had been on a HFD for 10 weeks. The HFD-induced obese group showed an additional continuous increase in body weight (BW) during the extended HFD period of 16 weeks, while SB treatment in HFD-fed mice (HFD+SB) suppressed the increase in BW (Fig. 3B). In addition, both blood glucose and serum insulin levels were lower after 16 weeks of SB treatment compared with those in non-treated HFD-induced obese mice (Figs. 3C, 3D). The dietary intake was not influenced by SB treatment (data not shown).
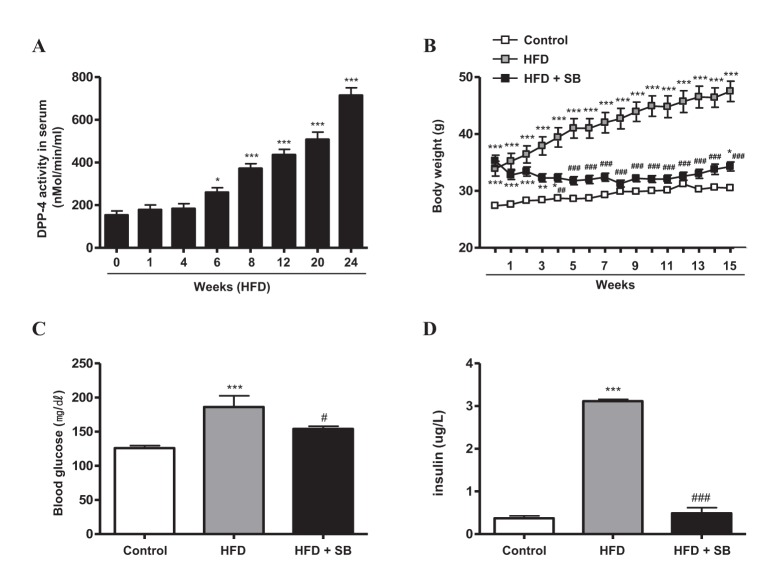
Fig. 3
The effects of SB on DPP-4 activity (A), body weight (B), blood glucose (C), and insulin secretion (D) in HFD-induced obese mice.
C57BL/6J mice were fed NCD (Control, n=9) or 60% high-fat diet (HFD) for 10 weeks. Mice in the control group were fed NCD for the additional 16-week period (Control, n=9). HFD-induced obese mice were divided into two groups; the mice were kept on HFD with SB incorporated at 5% wt/wt (HFD+SB, n=9) or HFD only (HFD, n=9) for the additional 16-week period. Data represent means±S.E. *p<0.05, **p<0.01, and ***p<0.001 vs. Control group; #p<0.05, ##p<0.01, and ###p<0.001 vs. HFD group.
![]()
Additionally, we evaluated the effects of SB on serum DPP-4 activity and mRNA expression levels of DPP-4, TNF-α, IL-1β, IL-6, and MCP-1 in peripheral blood cells (PBCs) from the experimental mice. While DPP-4 mRNA expression in PBCs was not different between the experimental groups, increased serum DPP-4 activity in HFD-induced obese mice was suppressed by SB treatment (Figs. 4A, 4B). In addition, mRNA expressions of TNF-α, IL-1β, IL-6, and MCP-1 in PBCs were down-regulated by SB treatment in HFD-fed mice (Figs. 4C–4F). Among the three cytokines that we measured in the serum obtained from the experimental groups (Figs. 4G–4I), only the IL-1β level was significantly increased by HFD feeding, and SB treatment significantly suppressed the increase in serum IL-1β (Fig. 4H).
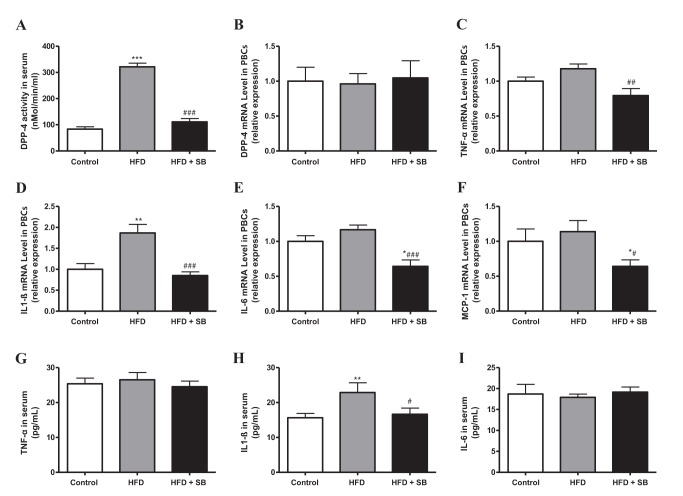
Fig. 4
The effects of SB on DPP-4 regulation (A, B), and TNF-α (C, G), IL1β (D, H), IL-6 (E, I), and MCP-1 (F) production in peripheral blood cells (PBCs) or serum of HFD-induced obese mice (as in Fig. 3).
At 16 weeks after combination with SB in HFD-induced obese mice, the mice were sacrificed and the PBCs were collected (B~F). Data represent means±S.E. *p<0.05, **p<0.01, and ***p<0.001 vs. Control group; #p<0.05, ##p<0.01, and ###p<0.001 vs. HFD group.
![]()
SB has different effects on DPP-4 and cytokines in the liver and kidney from HFD-induced obese mice
Among tissues, liver and kidney were reported to have a very high DPP-4 activity [1516]. Therefore, after 16 weeks of SB treatment in HFD-induced obese mice, we investigated the effects of SB treatment on mRNA expression of DPP-4 and cytokines and DPP-4 activity in liver (Figs. 5A–5F) and kidney (Figs. 5G–5L) obtained from the three experimental groups. Contrary to serum, DPP-4 activity and mRNA expression in the liver were not affected by SB treatment in HFD-induced obese mice. However, the treatment of HFD-induced obese mice with SB suppressed the increase in hepatic mRNA levels of TNF-α (Fig. 5C) and MCP-1 (Fig. 5F).
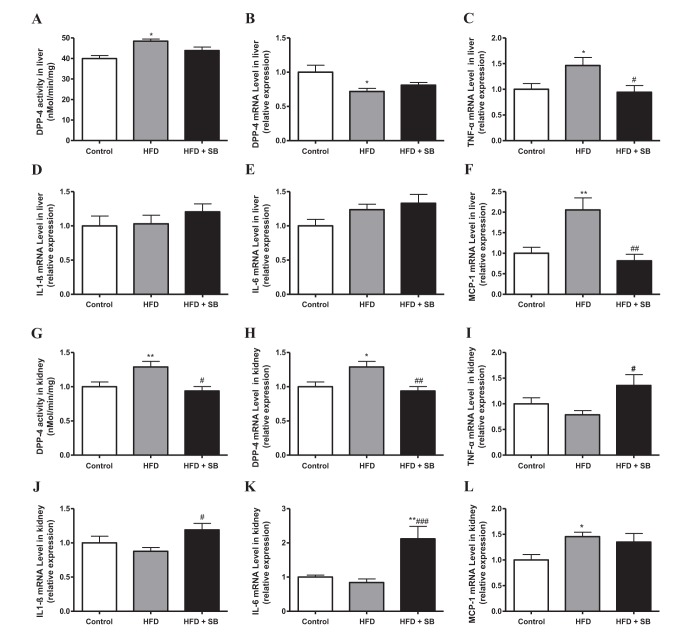
Fig. 5
The effects of SB on DPP-4 regulation (A, B, G, H), and TNF-α (C, I), IL-1β (D, J), IL-6 (E, K), and MCP-1 (F, L) production in the liver or kidney tissues of HFD-induced obese mice (as in Fig. 3).
Data represent means±S.E. *p<0.05, and **p<0.01 vs. Control group; #p<0.05, ##p<0.01, and ###p<0.001 vs. HFD group.
![]()
In the kidney (Figs. 5G–5L), DPP-4 activity and mRNA expression were increased in the HFD group compared with the control group, and these increases were suppressed in the HFD+SB group (Figs. 5G, 5H). However, to our surprise, the renal mRNA expression of TNF-α, IL-1β, and IL-6 was significantly increased by SB treatment in HFD-fed mice (Figs. 5I–5L).
Considering the regulating effect of SB on DPP-4 we measured the plasma active GLP-1 level in the experimental mice; however, plasma active GLP-1 levels were comparable between the groups (data not shown).
SB upregulates DPP-4 and increases cytokine secretion in cultured mouse mesangial cells
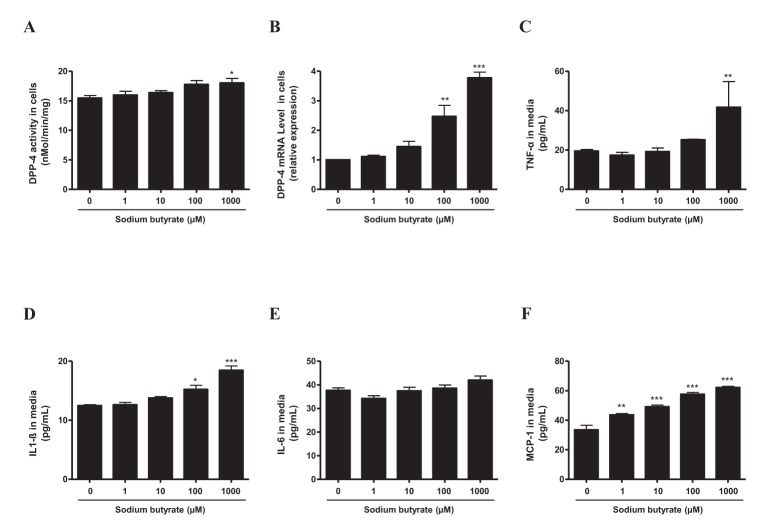
Finally, to better characterize different effects of SB depending on the concentration and tissue, an additional experiment was conducted using a mesangial cell line. Mesangial cells were treated with indicated concentrations (0~1,000 µM) of SB and incubated for 3 days. First, DPP-4 activity and mRNA level were increased by SB at 100 and 1,000 µM (Figs. 6A, 6B). Furthermore, secretion of TNF-α, IL-1β, and MCP-1 from cultured mesangial cells was increased significantly at high concentration of SB, whereas IL-6 production was not affected (Figs. 6C–6F).
DISCUSSION
DPP-4 is well known for its pivotal role in regulation of glycemia via catabolism of incretin peptides, and it plays many other roles in cell-matrix interactions and in the regulation of functional peptides [1617]. DPP-4 expression has been shown to be influenced by hypoxia, with hypoxia-inducible factor-1α (HIF-1α) being a strong inducer of DPP-4 gene and protein [18]. Besides HIF-1α, hepatocyte nuclear factor-1α (HNF-1α), interferons, retinoic acid, various cytokines, and advanced glycation end products have been shown to activate DPP-4 [1920212223242526]. Therefore, DPP-4 has implications for a potential pathophysiological role not only in metabolic disorders, but also in other disorders, including inflammatory diseases. Thus, further studies on factors regulating in vivo DPP-4 activity in both endogenous and exogenous manners seem necessary.
SB, being also known as an HDAC inhibitor, has been receiving much attention because of its wide range of metabolic actions and anti-inflammatory and anti-cancer effects [142728293031323334]. Although a previous study demonstrated that SB at a high concentration largely increases DPP-4 activity in HepG2 cells, it was beyond the physiologic range [8]. Other studies revealed that the responsiveness of different cancer cell lines to SB varied greatly. Especially, Jiang et al. [12] reported that while a low concentration of SB promoted cell proliferation and suppressed p21 Cip1 expression, a high concentration of SB induced growth arrest and cell apoptosis, accompanied by p21 Cip1 up-regulation and poly (ADP-ribose) polymerase cleavage. However, the study showed that HDAC inhibition by SB in cancer cell lines including HepG2 cells was clearly dose-dependent from low to high concentrations of SB, indicating differential effects of SB on between HDAC inhibition and the regulation of cellular growth.
Metabolic actions of SB have recently been revealed intensively [11283032]; however, it needs to be investigated further with regard to DPP-4. The present study evaluated not only the effect of SB on DPP-4 activity in vitro and in vivo, but also metabolic factors affecting DPP-4 activity. We found that SB suppressed DPP-4 activity at concentrations of 1~100 µM in a concentration-dependent manner, but the DPP-4 activity was increased at a high concentration of SB (1,000 µM). It has already been reported that a high level of SB (5 mM) increased DPP-4 activity in HepG2 cells [17]; however, there is no report suggesting that a low level of SB decreases DPP-4 activity. Previous studies revealed that SB has a suppressive effect on HDACs consistently [1231]. We also observed that SB increased Acetyl-H3 in a dose-dependent manner, in contrast to its effect on DPP-4. Thus, our study showed that DPP-4 has dual effects on DPP-4 activity in HepG2 cells, but not on HDAC activity. Our results are in line with a previous report, which showed that distinct dual effects of SB on cell growth did not match its inhibitory effect on HDAC in hepatic cancer cell lines [12].
We also evaluated the effect of high glucose on DPP-4 activity in HepG2 cells. Long-term exposure of HepG2 cells under the high glucose condition led to a significant increase in DPP-4 activity in the cells. Interestingly enough, in contrast to the results under a relatively low glucose condition (11 mM), under the high glucose condition, SB treatment of HepG2 cells with SB at low concentration (10~100 µM) did not lower, but increased DPP-4 activity, even earlier than high glucose condition only. Thus, SB has different effects on cells in context-dependent manners.
We also evaluated HFD on serum DPP-4 activity in mice. Serum DPP-4 activity was increased continuously from 6 weeks of HFD. This is the first study to show the time course of serum DPP-4 activity in HFD-induced animal model for a long-term period. In our previous study, we showed that serum soluble DPP-4 and DPP-4 activity were increased in patients with T2DM [5], while other researchers reported that lower DPP-4 levels following exercise training plus weight loss were related to increased insulin sensitivity in adults with metabolic syndrome [335]. Thus, our results and others indicate that metabolic dysregulation itself lead to an increase in serum DPP-4 activity which is actually the therapeutic target of diabetes for incretin effects.
Butyrate increased mitochondrial respiration at the cellular level, through the regulation of energy metabolism-related genes including peroxisome proliferator-activated receptor (PPAR), PPARγ coactivator-1, and carnitine palmitoyltransferase 1b at the molecular level [113637]. In addition, mice group fed a HFD including 5% SB showed body weight maintenance, muscle increment and blood glucose decrement [11]. To the best of our knowledge, there is no report about SB with respect to the in vivo regulation of DPP-4 activity. Therefore, to address this issue, we applied the same diet condition (60% HFD containing 5% SB) as the study performed by Gao et al. [11], and we obtained similar results, especially with respect to body weight and blood glucose. In addition, serum DPP-4 activity in HFD-induced obese mice was significantly suppressed in the group that received 5% SB mixed in the HFD. These results suggested that SB may be effective in preventing obesity and insulin resistance, with one of mechanisms being the regulation of DPP-4 activity. However, this suggestion was not supported by GLP-1 measurement data, which showed no difference in plasma active GLP-1 levels between the groups in the present study. SCFAs, specifically acetate and propionate, but not butyrate, has been shown to increase GLP-1 secretion, probably via a SCFA receptor free fatty acid receptor 2 (FFAR2), in experiments using colonic cell culture system and cecal SCFA infusion to mice [3839]. However, orally ingested SCFAs are absorbed in the small intestine and do not reach the cecum. Thus, oral administration of SCFAs was not shown to increase plasm GLP-1 level in mice [38]. If butyrate inhibits DPP-4 activity in whole body at a range of its physiologic concentrations, one would expect an elevation of circulating active GLP-1 level in SB-treated mice. However, as we observed, tissue- and context-dependent actions of SB might affect the results in the present study. Further studies are required on this issue.
The liver and kidney have significantly higher DPP-4 activities [3]. In addition, the main source of endogenous DPP-4 has been reported to be micro-vascular endothelial cells of some tissues such as liver and kidney [3151640]. To investigate the metabolic regulatory action of SB, we also measured on the regulation of AMP-activated protein kinase (AMPK), peroxisome proliferator-activated receptor gamma coactivator (PGC)1-α, and PGC1 expression in the liver tissues of HFD-induced obese mice. AMPK plays a key role in hepatic lipid metabolism, and it is generally known that high-fat diets reduce AMPK activity in the liver tissue [4142]. However, interestingly, our previous study showed that the phosphorylation of AMPK increased in the liver tissues of HFD-induced obese mice [43]. In the present study, the phosphorylation of AMPK was also significantly increased in the liver tissue of HFD-induced obese mice. And SB treatment in HFD-fed mice (HFD+SB) was increasingly induced the phosphorylation of AMPK more than HFD-fed mice group (Supplementary Fig. 1A). Furthermore, PGC1-α, and PGC1 expression were increased by SB treatment in the liver tissues of HFD-induced obese mice (Supplementary Fig. 1B). Other previous studies also indicated that PGC1-AMPK signaling contributes to improve the metabolic effects [4445]. Therefore, these results suggested that the regulation of AMPK, PGC1-α, and PGC1 expression might be shown a major metabolic function of SB. In addition, DPP-4 inhibitor may have renoprotective effects on diabetic kidney disease, regardless of its glucose-lowering effects [46]. In this study, SB treatment of HFD-induced obese mice decreased DPP-4 activity and mRNA expression in the kidney, whereas renal TNF-α, IL-1β, and IL-6 mRNA levels were increased. Therefore, we also checked the effects of SB using a mesangial cell line. In mesangial cells, the dual effects of SB on DPP-4 activity were not observed, but DPP-4 activity was increased by treatment with a high concentration of SB. Furthermore, SB treatment of this mesangial cell line increased DPP-4 mRNA expression and stimulated the secretion of several pro-inflammatory cytokines (TNF-α, IL-1β, and MCP-1) in a dose-dependent manner. Thus, although many studies have shown that SB has metabolically beneficial effects and anti-inflammatory effects [34], we found that SB has quite differentiated effects in the kidney for HFD-induced obese mice and mesangial cells. Hence, further studies on the context-dependent actions of SB are needed.
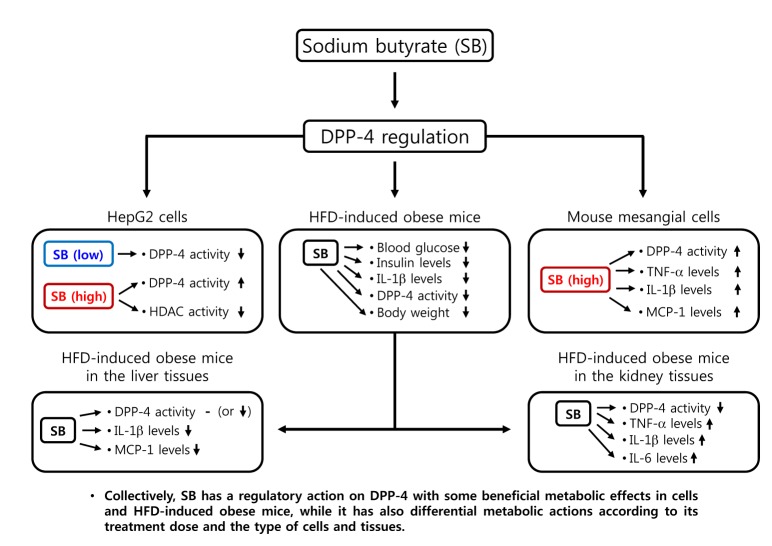
In conclusion, we demonstrated various effect of SB on the regulation of DPP-4 as well as other differential actions in HepG2 cells, mouse mesangial cells, and HFD-induced obese mice in context-dependent manners (Fig. 7). SB has a regulatory action on DPP-4 with some beneficial metabolic effects in cells and HFD-induced obese mice, while it has also differential metabolic actions according to its treatment dose and the type of cells and tissues. Therefore, further studies are required to evaluate its therapeutic relevance in metabolic diseases including diabetes and obesity and to determine safe and effective dosing of SB treatment for the metabolic diseases and their complications.
 XML Download
XML Download