

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
The anterior cingulate cortex (ACC), a region of the limbic system, is known for its role in functions such as cognition [12], decision making [3], memory [45], emotion [6], and pain [789]; however, the neural mechanisms underlying these functions have yet to be investigated. Emotion and pain processing in the ACC have been extensively studied through both imaging of the human brain and animal experiments. Human neuroimaging studies report that acute nociceptive stimuli activate several brain regions, including the ACC [1011121314]. Electrophysiology studies found that acute and chronic pain induce modifications in the synaptic plasticity of the ACC [1516171819], and inhibition of this plasticity produces consistent analgesic effects [7].
Optogenetics is a technique that is growing in popularity in the neuroscience field due to its ability to modulate the activity of specific cell subtypes during various behaviors [2021]. This technique has also been used in pain studies to look at involvement of the sciatic nerve [22], locus coeruleus [23], medial prefrontal cortex [242526], and ACC [272829]. Studies looking at the medial prefrontal cortex have reported that activation of excitatory neurons or inhibition of GABAergic circuits in the prelimbic cortex alleviates pain-like behaviors [242526]. Conversely, inhibition of excitatory neurons or activation of inhibitory neurons in the ACC reduced pain-like behavior [2829].
In the present study, we adapt optogenetic techniques to perform the conditioned place preference (CPP) test. The goal of this study was to determine whether optogenetics is able to modulate the motivational and emotional valence of stimuli to confirm our previous work using the designed receptor exclusively activated by a designed drug (DREADD) system [29]. CaMKII-cre and PV-cre mice were used to selectively inhibit excitatory neurons or activate the PV-positive inhibitory neurons in the ACC during the conditioning period of the CPP test, respectively. Complete Freund's adjuvant (CFA) was injected into the hind paw of mice and their response was observed with and without modulation by optogenetics. We found that inhibition of excitatory neurons was able to induce a preference for the conditioned chamber; however, activation of PV-expressing inhibitory interneurons had no effect on preference. The DREADD system was expressed in CaMKII-cre mice, and excitatory pyramidal neurons in the ACC were inhibited. Chemogenetic inhibition was able to alleviate CFA-induced hyperalgesia, as seen during optogenetic modulation. These results suggest that ACC excitatory neurons are critical mediators of both nociception and emotion.
Go to : 
METHODS
Animals
Adult (aged 8~12 weeks) CaMKII-cre or PV-cre mice (Jackson Laboratory, USA) were used. The animals were housed under a 12:12-h light/dark cycle with food and water provided ad libitum. All the experiments were conducted in accordance with the policy and regulation for the care and use of laboratory animals approved by the Institutional Animal Care and Use Committee at the Seoul National University.
Stereotaxic virus injection and optic cannula implantation
Prior to the CPP test, either the AAV-EF1α-DIO-eNPHR3.0-EYFP construct or the AAV-EF1α-DIO-EYFP construct was injected into 6-week-old CaMKII-cre heterozygous male mice. The PV-cre hetero mice or their wild-type littermates (control) were also injected with AAV-EF1α-DIO-ChR2-EYFP. Prior to the electronic von-Frey test, either the AAV-EF1α-DIO-hM4Di-mCherry or the AAV-EF1α-DIO-mCherry construct was injected into 6-week-old CaMKII-cre heterozygous male and female mice. The mice were anesthetized by intraperitoneal injection of ketamine-xylazine (0.1 mg ketamine per gram body weight, 0.01 mg xylazine per gram body weight) and the head was fixed in a stereotactic apparatus (Kopf Instruments, USA). A small craniotomy was performed and five holes were drilled for the CPP test, while two holes were drilled for the electronic von Frey test. For the CPP test, the two holes were drilled bilaterally between the hippocampus and the cerebellum for screw implantation. The other three holes were drilled for the virus injection and optic cannula implantation in the ACC. In the electronic von Frey test, the two holes were drilled on the bilateral ACC virus injection only. The virus was delivered using a 10-µl syringe (Hamilton, USA) and a 30-gauge metal needle. The injection volume and flow rate (0.5 µl at 0.1 µl/min) were controlled by an injection pump (WPI, USA). The virus was injected into both sides of the ACC (anteroposterior [AP]+1.0 mm from bregma, mediolateral [ML]±0.35 mm, dorsoventral [DV]−2.0 mm). After injection, the needle was left for an additional 7 min and was then slowly removed. The optic cannula (self-production) was implanted between the virus injection sites (AP+1.0 mm from the bregma, ML+0 mm, DV−1.25 mm). The optic cannula was then fixed using dental cement (Vertex Dental, The Netherlands).
Behavior
Conditioned place preference (CPP, Fig. 1)
The CPP apparatus consisted of two compartments (20×20 cm2) of identical sizes and a narrow, 10 cm-wide corridor connecting them. The two compartments had differently patterned walls (stripe and circles) to allow the mice to discriminate between them.
One day before the CPP test, CFA (30 µl of 100% CFA, F5881, Sigma-Aldrich, USA) was injected into the hind paw of the mice.
One day after CFA injection, the mice were assessed for their innate compartment preference. The mice were placed randomly in one of the two compartments and allowed free access to both the compartments for 15 min.
One day after the pre-test, the mice were conditioned for 6 consecutive days in bright light (130 lux). On each day, the mice were presented with two conditioning trials separated by 3 hours: one with laser stimulation in the compartment that was least preferred, and the other without laser stimulation in the opposite compartment (biased CPP). Conditioning with laser stimulation was performed in the compartment for which mice showed less preference during the pre-test. The stimulation involved continuous stimulation with a laser of wavelength 593 nm at an intensity of 7~9 mW/cm2. A laser of wavelength 473 nm was used in 300 ms pulses with a 2 s inter-pulse interval and an intensity of 20~25 mW/cm2 at the fiber tip. During conditioning, the mice were placed in each compartment and were not allowed access to the other compartment for 30 min. We changed the order of conditioning with or without laser stimulation daily.
After 6 days of conditioning, the mice were assessed for changed preference for conditioning compartments. As in the pre-test, the mice were placed in one of the compartments randomly and allowed to explore any compartment during the post-test for 15 min. The body position of the mice during the pre-test and post-test was tracked by the EthoVision XT video tracking software (Noldus Information Technology, The Netherlands). The exploration time in the conditioned compartment was normalized to the time spent in the compartment with less preference (exploration time during post-test/exploration time during pre-test).
 | Fig. 1Experimental schematic of the conditioned place preference testThe CFA-injected mice were first freely exposed to the apparatus for 15 min (Day 1, Pre-test). Next, the mice were conditioned with light in the compartment with less preference, and without light in the compartment with more preference (Days 2~7, conditioning). After 6 days of conditioning, the mice were assessed for changed preference for the conditioning compartments (Day 8, Post-test).
|
Electronic von Frey test
The electronic von Frey test apparatus (Ugo Basile, Italy) was placed underneath the testing grid to assess the mechanical threshold of the hind paw. A transparent plastic chamber was placed on the grid.
On the first day of testing, the baseline withdrawal threshold of the hind paw to mechanical stimuli was determined, followed by CFA injection into the right hind paw of each mouse (10 µl of 50% v/v CFA in saline).
After 2~3 hours of acclimatization in the testing chamber, mice were acutely anesthetized using isoflurane, followed by intraperitoneal injection of clozapine-N-oxide (CNO; 10 mg/kg in saline, BML-NS105, Enzo Life Science, USA). The mice were placed into the testing chamber for an additional 40 min of acclimatization.
Mice were habituated in the testing chamber for at least 2 hours at 23~25℃. On the first day of testing (Day 0), the baseline withdrawal threshold of the paw to the electronic von Frey apparatus was determined, and CFA was injected into the plantar face of the right hind paw of each mouse. On the third day after the CFA injection (Day 3), the mice were placed again in the testing chamber, and CNO was administered followed by additional acclimatization (10~30 min). During the following 80 min, the withdrawal threshold to the electronic von Frey apparatus was assessed again. The mechanical threshold was measured 6 times in every mouse with an inter-trial interval (ITI) of 6 min. The maximum and minimum values were excluded from the calculation, and the average was calculated from the remaining values.
Immunohistochemistry
Within 2 h of CNO injection, the mice that completed the electronic von Frey test were acutely anesthetized with isoflurane, and transcardially perfused with saline, followed by perfusion with 4% paraformaldehyde (PFA) in a 10 mM phosphate buffered saline (PBS; pH 7.4) solution. Whole brains were extracted and incubated in 4% PFA solution at 4℃ overnight and transferred to 30% sucrose the next day. Brains were dehydrated in sucrose solution for 2 days at 4℃ in a dark room. After fixation, the brains were sliced in a frozen state with OCT compound. Sections through the ACC (approximately 1.5 mm rostral to the bregma) from each animal were selected for c-Fos immunohistochemistry and placed in 50% glycerol in PBS.
The collected brain slices were washed in PBS for 5 min, followed by pre-blocking with 5% normal goat serum for 1 h. The sectioned slices were incubated at 4℃ overnight with a rabbit polyclonal antibody against c-Fos (1:200; sc-52; Santa Cruz Biotechnology, USA) in a dark room. The sections were rinsed with PBS, incubated for 2 h in Alexa 488-conjugated goat anti-rabbit antibody (1:200; A-11034; Invitrogen) while covered, and washed in PBS. The sections were treated with DAPI solution (10 µg/ml, D1306, Thermo Scientific, USA) for 5 min, and washed with PBS. The sections were mounted on slide glass, air-dried, cover-slipped, and images were acquired using a confocal microscope (Leica Microsystems, Germany).
In order to check whether neuronal activity in the ACC influences pain behavior, we manually counted c-Fos-positive (c-Fos(+)) cells within the mCherry-positive (mCherry(+)) population in a blinded state, and calculated the relative ratio of c-Fos expression (c-Fos(+)/mCherry(+)) after confocal microscopy using 5× magnification.
Data analysis
All data are presented as mean±SEM. Statistical comparisons were made using the unpaired t-test and two-way ANOVA performed in GraphPad Prism5 and SigmaPlot 10. Repeated-measures two-way ANOVA with post hoc Bonferroni test was used to compare the effect of CNO administration on the results of the electronic von Frey test. In all cases, the statistical significance was indicated by *p<0.05, and ***p<0.001.
Go to :
RESULTS
Previous studies have shown that the optogenetic modulation of ACC was able to control pain-related behavior [2829]. However, whether this modulation was also able to affect the motivational or emotional valence was not addressed. In the present study, we performed the CPP test to determine efficacy of optogenetic modulation of neuronal activity in the ACC.
After the CPP experiment, we conducted the electronic von Frey test using DREADD technology to confirm our previous finding that excitatory neurons in the ACC modulate pain-like behavior.
Inhibition of excitatory neurons in the ACC induces place preference
CaMKII-cre mice were injected with AAV-DIO-EYFP or AAV-DIO-eNpHR in the ACC bilaterally, and the optic cannula was implanted between virus injection sites. In our previous study, optogenetic inhibition of CaMKII excitatory neurons in the ACC was able to alleviate CFA-induced mechanical hyperalgesia [29]. Light with a wavelength 593 nm was continuously provided during the “light on” period. Therefore, we also used continuous light stimulation in the compartment with less preference during the conditioning period.
We found no difference in the exploration time in the compartment with less preference on Day 1 (Pre-test) between the EYFP (31.62±1.74 s, n=10) and eNpHR (28.39±0.98 s; n=14) groups (p=0.0978, unpaired t-test, Fig. 2A, B). However, after 6 days of conditioning1 (Day 8), there was a significant difference between the EYFP (0.97±0.06, n=10) and eNpHR (1.25±0.10; n=14) groups (p=0.0441, unpaired t-test) (Post-test, Fig. 2C). These results indicate that inhibition of CaMKII excitatory neurons in the ACC of CFA-induced mice provokes positive motivational valence.
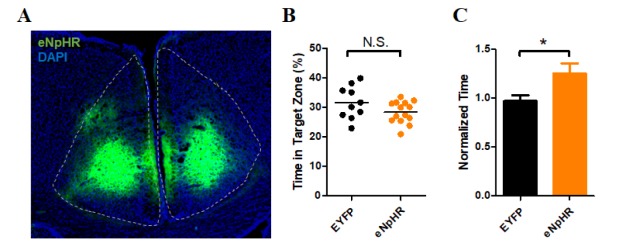 | Fig. 2Conditioned place preference in the CaMKII-eNpHR group.(A) Sample image of eNpHR expression in the ACC. (B) Result of exploration time in the less preferred compartment on Day 1 (Pre-test). There was no significant difference between the EYFP (31.62±1.74%, n=10) and eNpHR (28.39±0.98%; n=14) groups in exploration time (p=0.0978, unpaired t-test). (C) Result of the time spent in the conditioned compartment on Day 8 (Post-test, normalized to the pre-test exploration time). There was a significant difference between the EYFP (0.97±0.06, n=10) and eNpHR (1.25±0.10; n=14) groups (p=0.0441, unpaired t-test).
|
Activation of PV inhibitory neurons did not induce place preference
PV-cre (−) and PV-cre (+) mice were injected with AAV-DIO-ChR2 in the ACC bilaterally, and the optic cannula was implanted in between virus injection sites, as in the CaMKII-eNpHR experiments. Activation of PV-positive inhibitory neurons was able to alleviate CFA-induced mechanical hyperalgesia when light was provided at a frequency of 2 Hz during the “light on” period. The same light stimulation protocol was performed during conditioning.
The exploration times in the least preferred compartment on Day 1 (Pre-test) by the Cre(−) (29.50±3.23 s, n=4) and Cre(+) (27.11±1.90 s, n=8) groups were not significantly different (p=0.5110, unpaired t-test), as in the CaMKII-eNpHR experiments (Fig. 3A, B). However, we did not observe a significant difference between the Cre(−) (1.06±0.03, n=4) and Cre(+) (1.16±0.12, n=8) groups (p=0.7481, unpaired t-test) in the time spent in the conditioned compartment on Day 8 (Post-test, Fig. 3C). These results suggest that our stimulation protocol was not sufficient to induce positive motivation in chronic inflammation pain model mice.
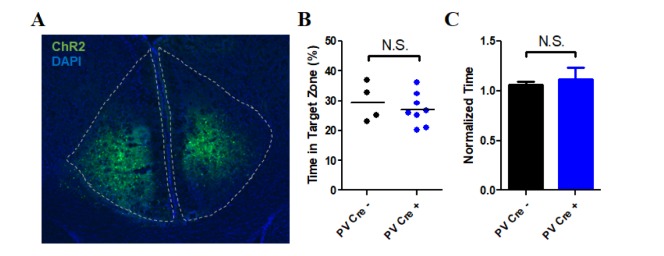 | Fig. 3Conditioned place preference in the PV-ChR2 group.(A) Sample image of ChR2 expression in the ACC. (B) Result of exploration time in the less preferred compartment on Day 1 (Pre-test). There was no significant difference between the cre(−) (29.50±3.23%, n=4) and cre(+) (27.11±1.90%, n=8) groups in exploration time (p=0.5110, unpaired t-test). (C) Result of the time spent in the conditioned compartment on Day 8 (Post-test, normalized to the pre-test exploration time). There was no significant difference between the cre(−) (1.06±0.03, n=4) and cre(+) (1.16±0.12, n=8) groups (p=0.7481, unpaired t-test).
|
Inhibition of excitatory neurons in the ACC modulates the paw withdrawal response
Previously, we showed that acute optogenetic inhibition of CaMKII excitatory neurons in the ACC diminished the CFA-induced mechanical hyperalgesia response. To demonstrate that chronic inhibition of excitatory neurons in the ACC is also able to alleviate mechanical hyperalgesia, we employed the DREADD system. CaMKII-cre mice were injected with AAV-EF1a-DIO-hM4Di-mCherry or AAV-EF1a-DIO-mCherry in the ACC bilaterally, and sutured for recovery. On Day 0, the baseline paw withdrawal threshold was measured, following which CFA was injected. On Day 3, after acclimatization to the test chamber, CNO was administered. The paw withdrawal mechanical threshold was reassessed after additional acclimatization during the next 80 min (Fig. 4A). Therefore, we chronically inhibited the neural activities of excitatory neurons in the ACC in hyperalgesia-induced mice.
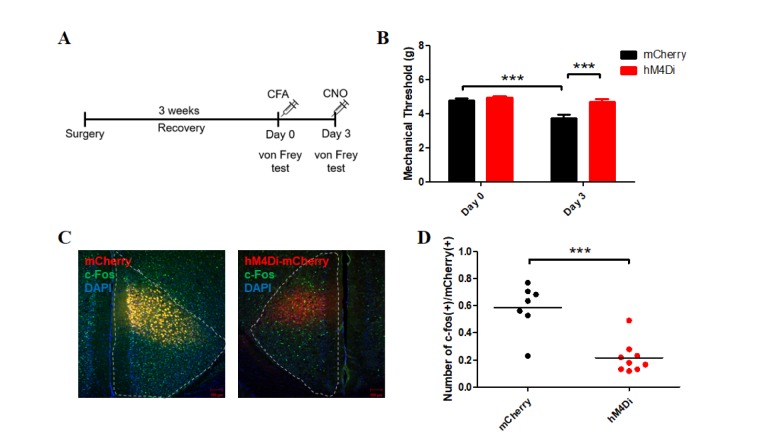 | Fig. 4Results of immunohistochemistry and electronic von Frey test in the CaMKII-hM4Di group.(A) Experimental schematic of the electronic von Frey test. Mice were allowed to recover from brain surgery for about 3 weeks. On Day 0, CFA was injected into the hind paw of each mouse, after determining the baseline response to the electronic von Frey apparatus. On Day 3, CNO was administered to each mouse 40 min before the experiment, and the response to the von Frey apparatus was measured. (B) Mechanical threshold responses to the electronic von Frey test on Days 0 and 3. There was no significant difference between the mCherry (4.81±0.09 g; n=20) and hM4Di (4.94±0.10 g; n=19) groups in the baseline responses (p=0.507, post hoc Bonferroni test) on Day 0. After CNO administration to both groups, the mechanical threshold showed a significant difference between groups (hM4Di [4.70±0.16; n=19], mCherry [3.76±0.19; n=20], p<0.001, post hoc Bonferroni test) on Day 3. The mCherry group displayed significantly different mechanical thresholds between days (Day 0 vs. Day 3, p<0.001). The mechanical threshold of the hM4Di group was not significantly different between days (Day 0 vs. Day 3, p=0.15). Repeated-measures two-way ANOVA with post hoc Bonferroni correction demonstrated a significant effect of CNO on Day 3 (p<0.001), significant effect of DREADD (p=0.002), significant difference between days (p <0.001), and a significant effect of interaction between two factors (Virus x Time, p=0.0013). (C) Images of the ACC of the mCherry (left) and hM4Di (right) groups after immunohistochemistry. (D) The ratio of activated neurons labeled by c-Fos among mCherry(+) neurons in the mCherry group was significantly different from that in the hM4Di group (p=0.0002, unpaired t-test).
|
There was no difference in the baseline mechanical paw withdrawal threshold between the mCherry (4.81±0.09 g; n=20) and hM4Di (4.94±0.10 g; n=19) groups (p=0.507, post hoc Bonferroni test) on Day 0 (Fig. 4B). CNO was administered in both the groups after induction of hyperalgesia, and while it restored the mechanical threshold in the hM4Di (4.70±0.16; n=19) group, it was unable to do so in the mCherry (3.76±0.19; n=20) group (p<0.001, post hoc Bonferroni test). Two-way ANOVA revealed that the effect of inhibitory DREADD was significant (p=0.002), and the interaction between virus and time was also significant (p=0.0013).
Immunohistochemistry results also supported this notion. CaMKII-positive neurons in the ACC were marked by red fluorescence by expression of mCherry either with or without the expression of the inhibitory receptor (hM4Di group and mCherry group, respectively) (Fig. 4C, D). The relative neuronal activities, based on c-Fos expression in the ACC, in the two groups were significantly different (p=0.0002, unpaired t-test), indicating that the administration of CNO successfully inhibited the excitatory activity of neurons during the electronic von Frey test.
Our results indicate that chemogenetic inhibition of excitatory neurons in the ACC was able to successfully alleviate CFA-induced hyperalgesia, as observed in previous optogenetic experiments.
Go to :
DISCUSSION
In the present study, we demonstrated that neurons in the ACC not only elicit nociceptive responses to painful stimuli, but also influence motivational state during pain. We found that inhibition of excitatory neurons in the ACC provided positive valence in mice with chronic inflammatory pain.
It has been suggested that pain relief could serve as a reward and induce a strong motivational state, and the corticolimbic projection is one of the circuits believed to be important in this function [3031]. In the previous study, optogenetic inhibition of CaMKII excitatory neurons in the ACC was able to alleviate mechanical hyperalgesia induced by CFA, and was rapidly reversible after the termination of illumination [29]. We here aimed to determine whether this effect could serve as a reward in painful situations. Therefore, in the current CPP protocol, the CFA-injected mice were conditioned with or without light for several days. We discovered that optogenetic inhibition of CaMKII excitatory neurons produced strong motivational to stay in the compartment where the light was provided.
The mechanical threshold test in our previous study revealed a similar alleviation of mechanical hyperalgesia through both the activation of PV-positive inhibitory neurons and the inhibition of excitatory neurons. However, the same result was not observed when PV-positive inhibitory neurons were activated. One possible explanation for this discrepancy could be the optogenetic stimulation protocol. The 20-Hz frequency light stimulation used to activate PV-positive inhibitory neurons might not have been as effective at inhibiting CaMKII-positive excitatory neurons as continuous light stimulation. Although this activation protocol was sufficient to affect pain-like behavior in CFA-injected mice, it might not be enough to induce alterations in emotional state. As mentioned above, the corticolimbic circuit from the ACC to NAc is believed to function as a reward in pain relief situations [31]. Different light stimulation frequencies applied to ACC inhibitory neurons resulted in different firing rates in ACC excitatory neurons [28]. However, inhibiting excitatory neurons continuously with eNpHR induce no firing during light stimulation. Therefore, these optogenetic modulations might have different effects on corticolimbic reward and motivation circuit activation. Additional stimulation protocols should be tested in future studies.
Another possible reason for this observed difference could be that we are activating additional types of behavior with continuous optogenetic stimulation. It has been shown that activation of PV interneurons in the ACC is involved in foraging decision-making, which could influence behavior outside of the pain relief situation [32]. Pain relief and reward area foraging effects could have been induced simultaneously to alter chamber preference in the CPP test.
The medial prefrontal cortex, especially the prelimbic cortex, is also a region recently studied for its role in pain perception [242526]. The results from optogenetic experiments indicate opposing roles for the prelimbic cortex and the ACC in both sensory and emotional pain-like behavior. There might be a connection between these regions [33] for them to induce these opposing effects; however, no study in the field of pain research has examined these two regions together. This could also be a topic for future studies.
In conclusion, we identified a role of excitatory neurons within the ACC in painful situations, and these results raise the possibility of this pathway as a target for effective treatment of painful states.
Go to :
 XML Download
XML Download