

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Respiratory tract is a primary target because it is the first barrier to inhaled dust particles, which are associated with increased respiratory morbidity and mortality. Available evidence indicates dust particles mediate airway inflammation, the progression of asthmatic diseases [1], pneumonia [23], and inflammation by pathogens, and that they are even linked to brain structural changes and stroke [45]. Lungs provide large surface areas for the exchange of gases, but in so doing are exposed to exogenous particles. Particle size critically determines inhaled particulate uptake [6]. Fine particulates (less than 10 µm) reach the alveolar region and are cleared slowly because of the lack of a clearing mechanism [789]. For this reason, smaller particles and its extended deposition can be used to understand related downstream responses. Fine dust particles prefer respiratory invasion of not only nasal and bronchial epithelial layers but also of deep alveoli and other extra-pulmonary compartments including fibrotic tissue, lymph nodes, and the bloodstream [1011].
Pulmonary health is also threatened by dust particles that induce inflammatory and allergic reactions and cause Ca2+ overloading and the production of reactive oxygen species (ROS) [12]. Dust particles induce inflammatory reactions by inducing the productions of cytokines and chemokines, such as, transforming growth factor β1 (TGF-β1), interleukin-1, IL-6, and IL-8 [13141516]. In addition, particle-induced oxidative stress triggers immune reactions in alveolar macrophages and lung tissues [13171819], and these reactions have been associated with Ca2+, a critical pro-inflammatory mediator signal [19]. Intracellular Ca2+ is known to act as a pivotal intracellular messenger, and has been demonstrated to regulate several pulmonary functions, such as, ciliary beat frequency and mucus secretion [20212223], and mechanical movement of fibroblasts [2425].
Dust particles are known to increase an intracellular Ca2+ concentration ([Ca2+]i), which up-regulates transcription factors and the productions of their downstream mediators in airway epithelial cells [19]. Therefore, dust particles may influence the developments and exacerbations of respiratory diseases directly or indirectly [14]. However, it cannot be ruled out the possible mechanism sustained dust particles is a regulatory factors for development of inflammatory signaling in respiratory fibroblasts.
Although the above-mentioned studies have explored Ca2+ response in airway epithelial cells exposed to several particle types from ambient or chemical sources, the relationship between calcium response and signaling proteins has not been previously examined in bronchial fibroblasts. Accordingly, the goals of this study were to define changes in intracellular Ca2+ and signaling mechanisms in lung fibroblast cells exposed to dust particles PM10 and to explore the therapeutic agents responsible for particles-mediated signaling mechanism.
METHODS
Reagents and cell culture
Human lung fibroblast cell line MRC5 cells were purchased from American Type Culture Collection (Rockville, MD). Fura-2-AM was purchased from Teflabs (Austin, TX). U73122 and its inactive analog, U73343 were from Tocris (Minneapolis, MN). Caffeine, clotrimazole (CLZ), 3-Aminobenzamide (3-AB), N-(p-amylcinnamoyl) anthranilic acid (ACA), 2-aminoethoxydiphenyl borate (2-APB), Chlorpromazine (CLP), N-acetylcysteine (NAC), lanthanum chloride (LaCl3), and all other chemicals were from Sigma. Dulbecco's Modified Eagle's Medium (DMEM), penicillin-streptomycin, trypsin-ethylenediaminetetraacetic acid (EDTA), 5-(and-6)-choloromethyl-2′,7′-dicholorodihydrofluorescin diacetate (CM-H2DCFDA), phosphate-buffered saline (PBS), fetal bovine serum (FBS), Pluronic F-127 (20% in DMSO), and 1,2-bis (2-aminophenoxy) ethane-N,N,N',N'-tetraacetic acid tetrakis, acetoxymethyl ester (BAPTA,AM) were from Invitrogen (Carlsbad, CA). MRC5 cells were incubated at 37℃ in a humidified 5% CO2/95% air atmosphere in DMEM containing 10% FBS, 100 U/mL penicillin, and 100 µg/mL streptomycin. When the cell culture reached 80% confluence, cells were dispersed by adding trypsin-EDTA for 2 min and then transferred to new culture dishes or to glass coverslip-covered dishes for Ca2+ measurements.
Asian dust particles (PM10) sampling, analysis, and measurement of particles size
Ambient air samples were collected in Incheon City, South Korea and analyzed as described previously [2]. Collected outdoor dust particles were sonicated for 3 min at maximal watt and sieved through filters at 10 µm in size (Mitex membrane filters, Millipore). PM10 suspensions consisted of: 48% SiO2, 12% AL2O3, 5% Fe2O3, 5% CaO, 4% K2O, 2.37% MgO, 2% Na2O, and 1% TiO2. PM10 suspensions were heated at 360℃ for 30 min to remove adhered microorganisms or other organic materials and stored at −20℃ until use. No LPS in heated PM10 was detected (below range of 0.005 EU/ml) by a Limulus amebocyte lysate assay kit (BioWhittaker, MD). Particles were diluted in water and its size was measured in a Zeta potential and particle size analyzer (ELSZ-1000, Otsuka Electronics, Japan).
Measurement of intracellular Ca2+ concentration ([Ca2+]i)
MRC5 cells cultured on cover glasses were incubated with 4 µM fura-2, AM in the presence of 0.05% Pluronic F-127 for 15 min in physiological salt solution (PSS) at room temperature in the dark, and then washed for 10 min with PSS (containing in mM: 10 D-glucose, 140 NaCl, 5 KCl, 1 MgCl2, 1 CaCl2, and 10 HEPES and titrated to pH 7.4). For the 0 Ca2+ extracellular solution, CaCl2 was replaced with 10 mM EGTA. Changes in intracellular Ca2+ concentrations were determined by measuring the fluorescence intensities using dual excitation wavelengths (340 and 380 nm) and an emission wavelength of 510 nm. Results are presented as fluorescence (F) ratios (Ratio=F340/380). Emitted fluorescence was monitored using a CCD camera (Photometrics, AZ) attached to an inverted microscope (Olympus, Japan) and analyzed with a MetaFlour system (Molecular Devices, PA). Fluorescence images were obtained at 1 sec intervals. Background fluorescence at each excitation wavelength was subtracted from raw signals.
Imaging of reactive oxygen species (ROS)
MRC5 cells grown on a cover glass in the presence or absence of PM10 were incubated in PSS containing the ROS fluorescence probe 10 µg/mL H2DCFDA for 5 min and then washed with PBS for 5 min at room temperature. Fluorescence images were obtained using a confocal laser-scanning microscope (Leica, Buffalo, NY) using an excitation wavelength of 488 nm for DCF and measuring emitted light intensities at 525 nm. DCF fluorescence was measured in five different regions of selected images and signals were normalized versus baseline. Images were obtained at 10 min intervals.
RESULTS
PM10-induced [Ca2+]i signal by extracellular Ca2+ in human lung fibroblast MRC5 cells
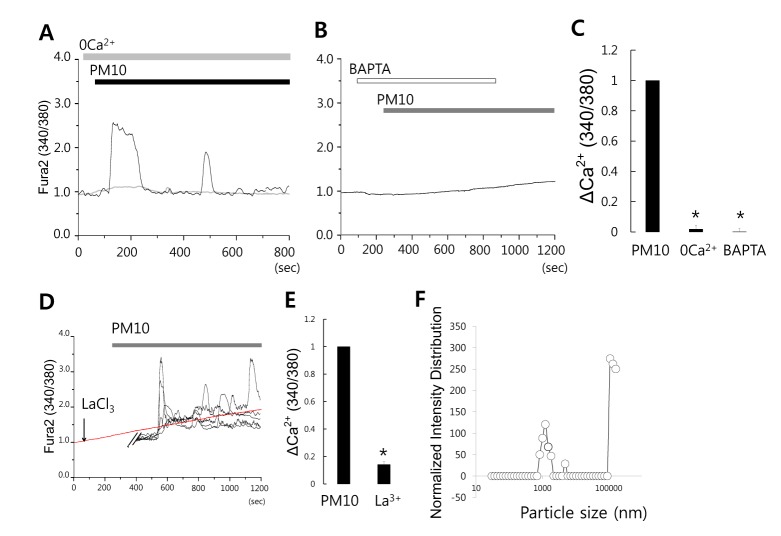
To characterize the source of Ca2+ induced by PM10, MRC5 cells were stimulated by PM10 in the presence or absence of extracellular Ca2+. The PM10-induced [Ca2+]i signal was dramatically reduced in the absence of extracellular Ca2+ (Fig. 1A and 1C; n=25 cells). When treated with BAPTA,AM as a broad range of Ca2+ chelator, the PM10-induced [Ca2+]i signal was blocked by the chelation of basal and increased Ca2+ (Fig. 1B and 1C; n=28 cells). These observations showed that PM10 induced an increased [Ca2+]i signal which was mediated by extracellular Ca2+ in MRC5 cells. To confirm PM10-induced [Ca2+]i increases were mainly mediated by extracellular media, cells were treated with PM10 in the presence of a non-selective Ca2+ channel blocker La3+. PM10-mediated [Ca2+]i increases were blocked by La3+ (Fig. 1D and 1E, n=60 cells). Collected dust particles were characterized as PM10 (less than 10 µm), which including less than 2.5 µm dust particles PM2.5 (Fig. 1F).
PM10-induced [Ca2+]i signal is dependent on the PLC/IP3 receptor pathway
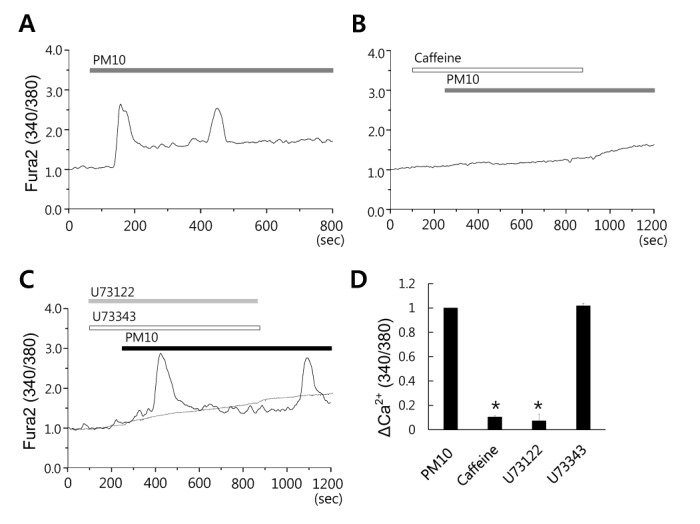
To determine the involvement of the phospholipase C (PLC)/IP3 receptor (IP3R) pathway in PM10-induced [Ca2+]i responses, MRC5 cells were pre-treated with caffeine, which is an antagonist of IP3 receptor (IP3R) in many types of cells [262728]. The PM10-induced [Ca2+]i signal was measured in pretreatment with 20 mM caffeine for 3 min. Caffeine suppressed the [Ca2+]i increase induced by PM10 (Fig. 2A and B, n=64 and 97 cells, respectively). To determine the role of PLC in the effect of PM10, cells were pretreated with U73122 (a specific blocker of PLC) or with its inactive analog U73343. U73122 blocked PM10-induced [Ca2+]i increases, but U73343 did not (Fig. 2B and 2C, n=31 and 60 cells, respectively). These results suggest that PM10-induced [Ca2+]i increases are dependent on the PLC/IP3R pathway in MRC5 cells.
PM10-induced [Ca2+]i signal is attenuated by inhibition of oxidative pathways
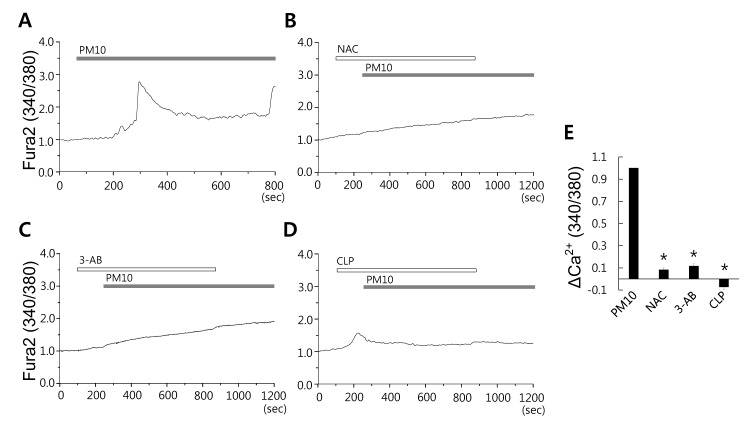
ROS generation by airway epithelia is a characteristic feature of respiratory diseases, such as, chronic obstructive pulmonary disease (COPD) [29], and therefore, the targeting of ROS represents a potential therapeutic strategy in such patients. Furthermore, dust particles have been shown to promote the production of ROS in bronchial epithelial cells [13]. To probe this situation we treated with PM10 in MRC5 cells in the presence of several ROS scavengers. PM10-induced [Ca2+]i signals were modestly attenuated by the antioxidant NAC (Fig. 3A and 3B, n=45 and 65 cells, respectively), indicating that ROS production is involved in Ca2+ signaling mechanism. It is known intracellular ROS increases polyADP-ribose (ADPR) polymerase 1 (PARP-1) activity, that this up-regulation is associated with subsequent DNA damage. PARP is a cardinal factor downstream of ROS production facilitates ADPR synthesis [30]. To determine whether PM10-induced ROS responses influence PARP-1 signaling, we used a PARP-1 inhibitor 3-AB [31]. Pretreatment with 3-AB was found to markedly inhibit PM10-mediated [Ca2+]i increases (Fig. 3C, n=36 cells). Cells were also pretreated with a Ca2+-CaM inhibitor CLP to determine whether CaM was involved in PM10-triggered [Ca2+]i signaling [32]. PM10-induced [Ca2+]i increases were inhibited by CLP (Fig. 3D, n=30 cells), indicating PM10-induced increases in [Ca2+]i in lung fibroblast cells are mediated by oxidative signaling and are dependent on PARP-1 signaling.
PM10-induced [Ca2+]i signal is required for the involvement of TRPM2 activation
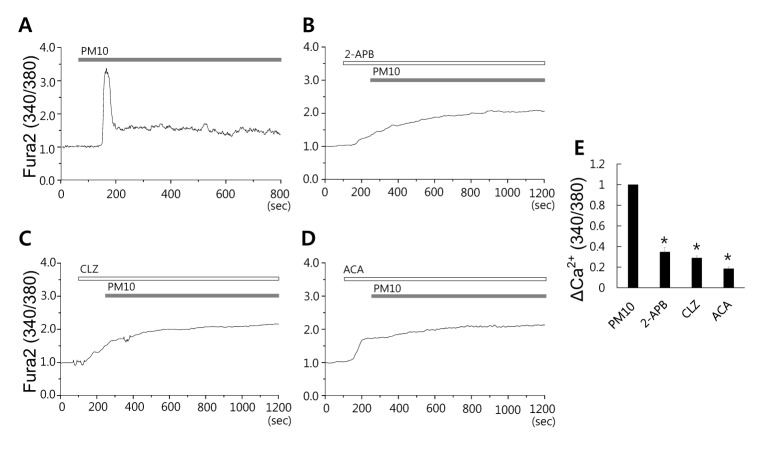
Increased PARP activity or [Ca2+]i increases are required for the activation of transient receptor potential melastatin 2 (TRPM2) [3334], and TRPM2 channel is activated by intracellular ADPR and ROS messengers, which leads to excessive Ca2+ influx [230]. To determine whether the PM10-induced [Ca2+]i signal was mediated by TRPM2, MRC5 cells were pretreated with several types of TRPM2 blockers 2-APB, CLZ, or ACA and then treated with PM10. It is known that 2-APB blocks ADPR-induced TRPM2 currents in several cell types [3536]. We found pretreatment with 2-APB or CLZ (Fig. 4A, 4B, and 4C, n=51, 32, and 60 cells, respectively) also significantly decreased PM10-induced Ca2+ influx into lung fibroblast cells. The pretreatment of derivative anthranilic acid, N-(p-amylcinnamoyl) anthranilic acid ACA [36] markedly decreased PM10-mediated influx despite initially increasing [Ca2+]i (Fig. 4D, n=52 cells). These results suggest that the [Ca2+]i response observed in PM10-stimulated bronchial epithelial cells required PARP/ADPR signaling to mediate activation of oxidative stress-dependent channels such as TRPM2 activation.
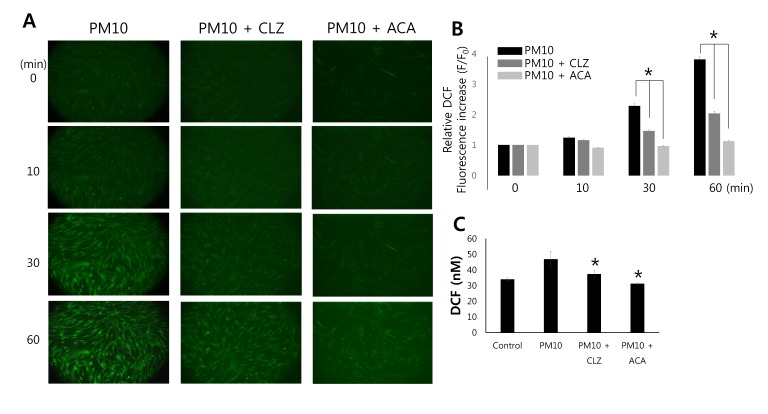
PM10 induced ROS production and ROS levels were reduced in the presence of TRPM2 blockers
MRC5 cells were loaded with H2DCFDA to determine extents of PM10-induced ROS signal. DCFDA fluorescence, which was used to detect H2O2, was time-dependently increased by PM10 (Fig. 5A; n=7), but pretreatment with both TRPM2 inhibitors CLZ or ACA resulted in the disappearance H2O2 from cells (Fig. 5B and 5C). To avoid the effect of dilution vehicle, PM10 suspension was included with DMSO. These result suggested that PM10 induced ROS signal with time-dependent manner and the ROS signal attenuated by both TRPM2 blockers, CLZ and ACA.
DISCUSSION
Ambient epidemiologic studies have demonstrated that increases in the morbidities and mortalities of pulmonary diseases are correlated with pollution of the environment by particulate matter (PM). Thus, clarifying the mechanisms of airway diseases induced by PM would support understanding of their pathogeneses. This study demonstrates that PM10 mediates ROS production and [Ca2+]i signaling in a human lung fibroblast cell line. In view of the inflammatory effects of dust particles on respiratory organs such as bronchial epithelia is not surprising; however, the present study shows inflammatory signaling upstream of PM is primarily associated with oxidative stress-dependent Ca2+ channels such as TRPM2, which triggers and promotes an inflammatory stream of extracellular Ca2+ into lung fibroblasts.
Ca2+ homeostasis is related to a variety of cellular functions, which include protein synthesis, transcription factor activation, cell migration, proliferation, and the transduction of extracellular signals into cells [37]. [Ca2+]i is tightly maintained by Ca2+ pump, which hydrolyzes ATP to provide the energy for Ca2+ movement and for store operated Ca2+ influx channels, such as, stromal interaction molecule 1 (STIM1) and Orai. Few studies have addressed the pathological aspects of Ca2+ signaling in pulmonary fibroblasts, although a large number of studies have been undertaken on non-pulmonary fibrotic organs, such as, heart [38], liver [39], and kidneys [40]. In the present study, PM10 evoked a delayed [Ca2+]i signal (Fig. 1), indicating the involvements of other signaling mechanisms prior to Ca2+ influx, and thus, it is important to determine whether dust particles induce ROS-mediated signaling. Although previous report addressed that air pollutants evoke both Ca2+-dependent or -independent mechanisms [19], our findings for the first time suggest that dust particle-mediated ROS production participates in [Ca2+]i signaling events via TRPM2 activation in pulmonary fibroblasts, which further suggests dust particles-mediated TRPM2 activation initiates inflammatory events such as increased proinflammatory cytokines level. On the other hands, TRPM2 activation, which enables excessive Ca2+ influx, phosphorylates and activates the PLCγ1/protein kinase Cα (PKCα) pathway, which subsequently induces airway epithelial tight junction disruption [41]. Particulate stimuli are common events in NLRP3 inflammasome activation [42].
We cannot exclude the possibility that additional [Ca2+]i signaling mechanisms are involved. Our results indicate dust particles elicit [Ca2+]i increases in a IP3/IP3R pathway dependent-manner. Kyung et al. [13] suggested that dust particles may have fibrotic potential in bronchial epithelial cells. This mechanism also allows us to predict and test of potential novel approaches for the treatment of pulmonary fibrotic diseases by long-term exposure of dust particles. However, the mechanism responsible for dysfunctional PLC/IP3/IP3R signaling by dust particles requires additional investigation in pulmonary fibrosis.
PM deposited in alveolar epithelia causes structural modifications in intracellular organelles and pathological changes in alveolar cells [43]. Furthermore, PM-mediated oxidative stresses are closely related to cell toxicity. In one study, PM2.5-mediated DNA damage was abolished by anti-oxidant enzymes or deferoxamine, indicating oxidative mediators are involved in the induction of DNA damage [44]. Oxidative events are also associated with many pathologic conditions, such as, pulmonary fibrosis and COPD [294546]. Indeed, blockade of ROS signaling or TGF-β1 has an anti-fibrotic effect in lung tissue [134647]. In addition, at elevated doses, PM appears to increase ROS levels and subsequent mitochondrial damage, such as mitochondrial swelling and vacuolation, in alveolar type II cells [43].
This study provides the role of [Ca2+]i signaling induced by dust particles in respiratory fibroblast cells. Currently, no therapeutic strategy targets [Ca2+]i signaling in fibroblast-mediated pulmonary diseases [48], but our results suggest potential means of controlling fibrotic pulmonary diseases by manipulating [Ca2+]i signaling events. In particular, we suggest antioxidant, [Ca2+]i signaling pathway inhibitor, and TRPM2 inhibitor treatments should be considered for therapeutic trials in fibrotic diseases. Importantly, this study shows the modulation of dust particles-mediated [Ca2+]i signaling has the potential to generate a new wave of therapies targeting pulmonary fibroblasts.
 XML Download
XML Download