

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Expression of K+ channels is developmentally regulated in cochlear outer hair cells (OHCs). Before hearing onset, delayed rectifier K+ current (IK,emb, IK,neo) [1], inward rectifier K+ current (IK1) [2], small conductance Ca2+ activated K+ current (ISK) [34], and K+ current activated at negative potentials (IK,n) are sequentially expressed in murine cochlear OHCs [5]. The reports, which suggest that the timely expression of K+ channels is important for the functional maturation of cochlear hair cells [156], raise a possibility that K+ channel expression in the early stage might be different in the cochlear hair cells of genetically abnormal animals, such as mouse models for human deafness.
We reported [7] the characteristics of K+ outward currents recorded in the OHCs of immature circling mouse (P0~P6), a mouse model for human deafness (DFNB6 type) [89], as a scanning electron microscope study demonstrated that the stereocilliary defects of OHCs were observable as early as P10 and were more severe in the OHCs than in the inner hair cells (IHCs) at P18 in circling mice [10].
This is a study following the previous study which demonstrated the subtle biophysical and pharmacological differences between circling mice (homozygous (cir/cir) mice) and their littermates (heterozygous (+/cir) mice). While investigating the pharmacological aspects of K+ outward currents in the previous study [7], we tested TEA and 4-AP only, not linopirdine, a known inhibitor of IK,n, as IK,n is not reported to develop before P6 in mice OHCs [5]. However, from succeeding experiments using linopirdine, we found significant inhibitory effects of linopirdine on K+ outward currents of P0~P6 heterozygous (+/cir) or homozygous (cir/cir) mice and we will discuss this finding in this report.
METHODS
Animal and cochlear preparation
Female heterozygous (+/cir) mice were mated with male homozygous (cir/cir) mice (circling mice), and their offspring were used in this study. The circling mice strain has been maintained for more than 10 generations by breeding between female heterozygous (+/cir) mice and their male siblings (homozygous (cir/cir) mice) at the Animal Facility of Dankook University since 2007. The data presented were obtained from pups between P0 and P6. Genotypes were assessed by polymerase chain reaction analysis according to our previous report [11]. The Dankook University Institutional Animal Care and Use Committee (DUIAC) approved this study.
After the mice received deep anesthesia with isoflurane, their cochleae were removed and dissected in an ice-cold solution composed of (in mM): NaCl (124), KCl (5), KH2PO4 (1.25), glucose (10), NaHCO3 (26), CaCl2 (2), MgSO4 (1.3), and sucrose (20). The pH was 7.4 when aerated with 95% O2 and 5% CO2 and the osmolarity was about 305 mOsm. After removing the bony part and modiolus, the dissected cochleae were transferred to a submersion-type chamber mounted on an upright microscope and immobilized under a nylon mesh fixed to a stainless steel ring. The chamber was perfused continuously with the same solution used during preparation.
Electrical recording
Whole cell currents were recorded from OHCs located in the middle turn of the cochlea. All experiments were performed at room temperature using an EPC-8 (HEKA, Lambrecht, Germany) amplifier. Electrodes (3~5 MΩ) were filled with a solution containing (in mM): K-gluconate (108), EGTA (0.6), KCl (5), HEPES (10), Na2GTP (0.3), MgATP (1), KOH (30), sucrose (47), and QX 314 (5). All chemicals except for QX 314 (Tocris) were purchased from Sigma Chemicals Co., unless otherwise stated. Except for fast capacitance cancellation at the cell-attached stage, series resistance was not compensated and no corrections were made for the liquid junction potentials. The data were filtered at 5 kHz (EPC-8, HEKA), digitized at 10 kHz, and stored in the computer via a home-made program (R-Clamp 1.23) for offline analysis. The stored data were analyzed using Clampfit 9.0 (Molecular Devices), Origin 7.0 (Origin Lab). Data were expressed as the mean±SEM. An independent t-test was used for comparisons. Null hypotheses of no difference were rejected if p values were <.05.
RESULTS
Characteristics of K+ currents expressed in OHCs during the first postnatal week
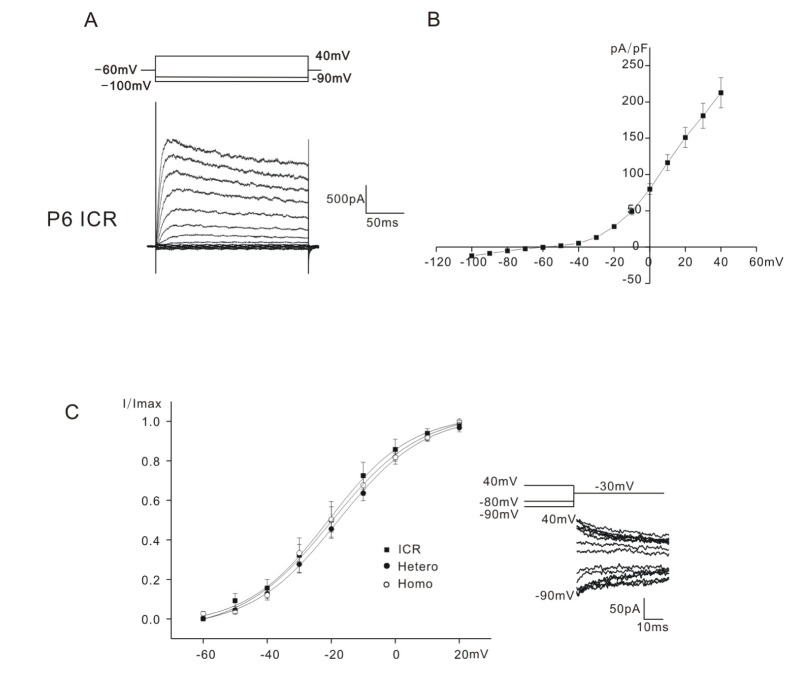
Depolarizing voltage steps from –100 mV to 40 mV (holding potential: –60 mV) evoked small inward (<100 pA) and slowly inactivating outward K+ currents in the OHCs of all mice tested. In Fig. 1A, only the data obtained from ICR mice was presented, as those for heterozygous (+/cir) and homozygous (cir/cir) were already presented in the previous paper [7]. All the K+ outward currents were similar to that reported previously (a delayed rectifier-type K+ current, IK,neo [1], for neonatal cells). The outward currents activated at potentials close to –50 mV in all mice tested. Small inward currents did not show any active current decay, which was usually observed in mice OHCs older than P9 [5] even at –120 mV (Data not shown). The current density-voltage relation of ICR mice is presented in Fig. 1B. The current density measured at 40 mV was 212.6±20.8 pA/pF in ICR mice (n=11). (In the previous paper [7], they were 227.1±18.9 pA/pF (n=16) in heterozygous (+/cir) mice and 238.9±14.3 pA/pF (n=22) in homozygous (cir/cir) mice).
Activation curves were derived from tail currents at –30 mV. The activation curves shown in Fig. 1C were obtained by plotting the normalized tail currents against different prepulse potentials from –90 mV to 40 mV. Data were fitted by a modified first-order Boltzmann equation: y=A2+(A1–A2)/{1+exp(V-Vhalf/S)} where y is the normalized peak current, A2 is the minimal normalized peak current, A1 is the maximal normalized peak current, Vhalf is the potential of half-maximal activation, V is the commanding potential, and S is the slope factor representing the voltage sensitivity of activation. Vhalf was –20.7±4.7 mV (n=7, ICR mice), –21.3±1.7 mV (n=9, heterozygous (+/cir) mice), and –20.8±3.3 mV (n=11, homozygous (cir/cir) mice). The slope factor was 9.8±0.9 mV (n=7, ICR mice), 13.8±1.8 (n=9, heterozygous (+/cir) mice) and 11.4±1.0 mV (n=11, homozygous (cir/cir) mice). They were not significantly different. In the previous paper, the reported values of Vhalfs were –8.1 (heterozygous (+/cir) mice) and –17.2 (homozygous (cir/cir) mice) [7]. In the previous paper, the activation curves were derived by calculating conductance (peak current/(Vcommand–Vholding)) at each commanding potential (Vholding=–60 mV), while in this paper, we used the tail current analysis. This might be the cause of the difference in Vhalfs.
Sensitivity of K+ currents to linopirdine
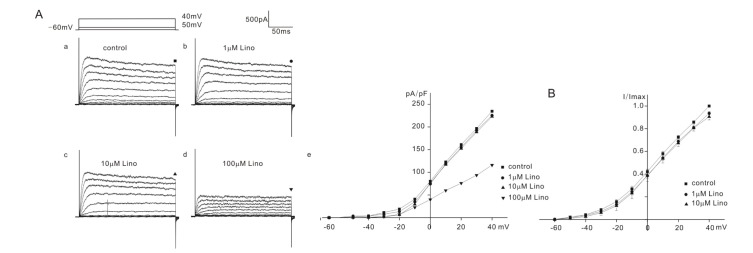
The above results indicated that biophysical properties of OHC K+ outward currents of heterozygous (+/cir), homozygous (cir/cir), and ICR mice were similar but quite different from those of IK,n reported in OHCs of older mice aged more than P9 [5] in which 1) hyperpolarizing voltage steps elicited only small inward K+ currents, 2) they did not show any active decay and 3) outward currents activated at potentials close to –50 mV, while they activated at potentials negative to –100 mV in OHCs older than P9 when IK,n developed fully, which suggested that K+ outward currents do not have the IK,n component. In spite of this, we tested the effects of linopirdine (1, 10, and 100 µM), a known blocker of IK,n to confirm that IK,n did not develop early in OHCs of homozygous (cir/cir) or heterozygous (+/cir) mice.
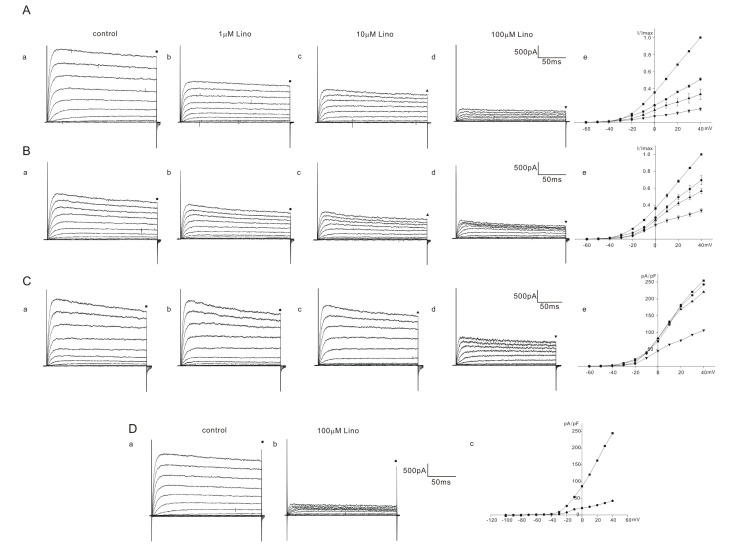
K+ currents were elicited by a series of voltage steps from –60 mV to 40 mV (holding potential: –60 mV) and the current amplitudes were measured at the end of the voltage steps (200 ms) for analysis. The responses of K+ outward currents were not simple but were categorized into 2 or 3 groups (severe, moderate, mild) according to the extent of inhibition by linopirdine in homozygous (cir/cir) or heterozygous (+/cir) mice. Among 13 cells of homozygous (cir/cir) mice, 5 cells showed a severe block, 4 cells showed a moderate block, and 4 cells showed a mild block. In 5 cells showing a severe block, the K+ current recorded at 40 mV was reduced to 51.0±1.6% (n=5), 33.1±6.1% (n=5), and 15.7±2.1% (n=5) of the control by 1 µM, 10 µM, and 100 µM linopirdine, respectively (Fig. 2A). In 4 cells showing a moderate block, the corresponding values were 69.5±5.3% (n=4), 56.7±3.0% (n=4), and 33.8±2.3% (n=4) of the control by 1 µM, 10 µM, and 100 µM linopirdine, respectively (Fig. 2B). Among 4 cells showing a mild block, we succeeded in observing a 100 µM response only in one cell. In the remaining 3 cells, we only observed the effects of linopirdine up to 10 µM. The K+ current at 40 mV was reduced to 91.7±4.7% (n=3, 1 µM) and 85.7±1.0% (n=3, 10 µM). The 100 µM response in one cell is shown in Fig. 2C. The currents at 40 mV were reduced to 90.5% (1 µM), 85.7% (10 µM), and 42.8% (100 µM). Linopirdine was not effective at hyperpolarizing voltages (–100 mV~–70 mV). Fig. 2D shows the representative data. No inhibition at the hyperpolarizing voltages was also observed in heterozygous (+/cir) and ICR mice (data not shown).
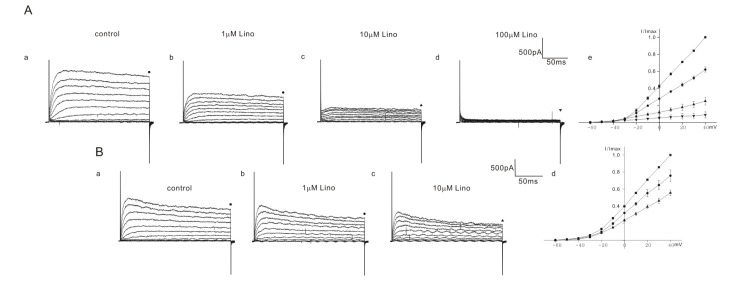
Among 10 cells of heterozygous (+/cir) mice, 4 cells showed a severe block and 6 cells showed a moderate block. In the OHCs of heterozygous (+/cir) mice showing a severe block, the current was reduced to 62.1±3.1% (n=4, 1 µM), 25.0±4.5% (n=4, 10 µM), and 8.9±3.5% (n=4, 100 µM) (Fig. 3A), while in the OHCs showing a moderate block, the values were 75.7±6.6% (n=6, 1 µM) and 55.4±3.4% (n=6, 10 µM) (Fig. 3B). We failed to record a 100 µM response in cells showing a moderate block.
In ICR mice, linopirdine responses were quite different from those in homozygous (cir/cir) or heterozygous (+/cir) mice. There were no cells showing a severe block caused by linopirdine among the total 9 cells recorded. We succeeded in observing a 100 µM response only in one cell among the 9 cells recorded. In the remaining 8 cells, responses only up to 10 µM were observed. In 8 cells, the currents were reduced to 93.6±1.4% (n=8, 1 µM) and 90.5±2.7% (n=8, 10 µM) (Fig. 4B). In one cell, the currents at 40 mV were reduced to 94.7% (1 µM), 92.1% (10 µM), and 50.4% (100 µM) (Fig. 4A).
We also checked the effects of linopirdine on small inward currents recorded by hyperpolarizing voltage steps (from –100 mV to –70 mV). However, we did not observe any inhibition in this voltage range in all cells tested (data not shown).
DISCUSSION
Currently, only two types of K+ currents, a transiently expressed inward rectifier (IK1) [2] and a delayed rectifier-type outward (IK,neo) [112], have been reported in P0~P6 mice OHCs. A small conductance Ca2+-activated K+ current (ISK)(>P6) [34] and a K+ current activated at negative potentials (IK..n) (>P8) are the next ones expected to develop in OHCs [5]. In this report, we showed the varying degree of inhibition of K+ currents by linopirdine, an inhibitor of members of the KCNQ family K+ channels [131415], in K+ outward currents of P0~P6 heterozygous (+/cir) or homozygous (cir/cir) mice OHCs. Although the IC50s values (1~10 µM, in cases of a severe block in homozygous (cir/cir) or heterozygous (+/cir) mice) fall within the reported range of values for the inhibition of KCNQ currents [141516], we do not think it likely that the K+ outward currents have some component of IK,n which develops earlier than expected in homozygous (cir/cir) or heterozygous (+/cir) mice OHCs, as the biophysical and pharmacological properties of K+ currents in this study do not match well with those of IK,n reported in guinea-pig [17] or mice OHCs [5]. The possibility of the involvement of ISK was ruled out in the previous study with respect to TEA sensitivity [7].
Firstly, unlike IK,n reported in guinea-pig [17] or mice [5], hyperpolarizing voltage steps elicited only small inward currents without active current decay in P0~P6 heterozygous (+/cir) and homozygous (cir/cir) mice. This small inward current (less than 100 pA at –100 mV) might be influenced by a holding potential of –60 mV. However, regardless of the holding potential, IK,n was easily observable in OHCs where IK,n was fully developed [51718]. The fact that linopirdine did not inhibit inward currents recorded at hyperpolarizing voltage steps also supports the claim that IK,n is not present in heterozygous (+/cir) or homozygous (cir/cir) mice OHCs.
Secondly, the activation curves obtained from homozygous (cir/cir) and heterozygous (+/cir) mice are similar to that obtained from P6 mice OHC in which K+ outward currents activate at potentials close to –50 mV, whereas they activate at potentials negative to –100 mV in P12 mice OHCs where IK,n is already developed [5].
Thirdly, K+ outward currents in homozygous (cir/cir) or heterozygous (+/cir) mice are sensitively blocked by TEA in the millimolar range [7]. We do not know the underlying ion channel subtypes of the K+ outward currents of heterozygous (+/cir) or homozygous (cir/cir) mice OHCs, but if it is KCNQ4 as suggested by Marcotti and Kros in developing mice OHCs [5], it is not in line with our results because KCNQ4 is known to be relatively insensitive to TEA [1920]. Moreover, the fact that linopirdine did not cause any further inhibition after K+ currents were fully inhibited by TEA (data not shown) in heterozygous (+/cir) or homozygous (cir/cir) mice also supports our notion.
Marcotti and Kros reported that linopirdine up to 200 µM was not effective at all in immature mice OHCs (<P6) [5]. Up to 10 µM linopirdine, we observed similar results in ICR mice OHCs; however, in one cell, we did observe that 100 µM linopirdine inhibited K+ current at 40 mV by 50%. In hippocampal neurons, it was reported that linopirdine exceeding 100 µM inhibited the delayed rectifier K+ currents by 50% [21]. Considering the non-specific blocking effects of linopirdine on other currents [2122], K+ current inhibition by a high concentration of linopirdine might not be surprising even in immature OHCs. Species difference should be considered.
In this study, we showed the varying degree of inhibition (severe, moderate, mild) by linopirdine. This indicates that developing OHCs are not homogeneous with respect to ion channel expression. We usually used cells located in the cochlear middle turn, but not always those in the same low and the same location. It is not clear whether this inhomogeneity stems from positional difference along the organ of Corti or other unknown factors.
The purpose of this study was to investigate the ion channels contributing to OHC degeneration starting from the second postnatal week in homozygous (cir/cir) mice [10]. We do not know the exact function of this linopirdine-sensitive K+ channel in the development of homozygous (cir/cir) mice OHCs, but it can be suggested that this K+ channel does not play a significant role in OHC degeneration because a similar K+ channel is also expressed early in heterozygous (+/cir) mice OHCs.
 XML Download
XML Download