

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Neuropathic pain is caused by damage to or pathological changes in the peripheral or central nervous system [1]. To date, the mechanisms of neuropathic pain are poorly understood; however, spontaneous pain, hyperalgesia, and allodynia have been recognized as cardinal signs. Animal models of neuropathic pain have been developed to simulate clinical settings such as causalgia[2]. Specifically, chronic constriction injury (CCI) [2], spinal nerve ligation (SNL) [3], and spared nerve injury (SNI) [4] models are used extensively to study the mechanisms of neuropathic pain. Neuropathic pain is also known to be intractable, where treatment using pharmacological drugs is associated with several side effects such as intolerance, addiction, and poor efficacy [5678]. Therefore, a new treatment strategy for treatment of neuropathic pain is necessary and transplantation of stem cells, a novel therapeutic modality developed for intractable neurological diseases, is a potential alternative.
Compared to stem cells of embryonic, bone marrow, and adipose origin, human umbilical cord blood-derived multipotentstem cells (hUCB-MSCs) are preferred for clinical application because they have a higher number of mesenchymal progenitor cells per unit volume and are less carcinogenic [91011]. We previously reported that subcutaneous injection of well-characterized hUCB-MSCs[1213] reduced severe pain in patients with Buerger's disease [14].
Matrix metalloproteinases (MMPs) are strongly implicated in inflammation and tissue remodeling associated with various neurodegenerative diseases, through cleavage of extracellular matrix proteins, cytokines, and chemokines[151617181920]. Recently, MMP2 has been shown to be crucial for maintenance of latephase neuropathic pain and an inhibitor of MMP2 was shown to effectively treat neuropathic pain in an SNL model [21]. In a preliminary study, we found that in vitro hUCB-MSCs produce large amounts of endogenous tissue inhibitor of MMP2 (TIMP-2).
The aim of this study is to evaluate if subcutaneous transplantation of the hUCB-MSCs can exert antinociceptive effects in vivo, using three different kinds of rat peripheral neuropathic pain models. Regarding pharmacological mechanisms of hUCB-MSCs for its antinociceptive effects in vivo, we studied theantinociceptive effects of the transplanted hUCB-MSCs onbiomolecular expression levels of MMP family and TIMP-2, an endogenous tissue inhibitor of MMP-2, and pain-related molecules such as c-fos, calcitonin gene-related peptide (CGRP), phosphorylated extracellular signal-related kinase (p-ERK), and phosphorylated p38 (p-p38).
Go to : 
METHODS
Animals, husbandry and ethical approval
Male Sprague-Dawley rats (Orient Bio Inc., Gyeonggido, Republic of Korea) weighing 150 to 200 g were used in this study, maintained in cages covered with soft bedding, with 3 animals per cage. The animals were divided into 2 groups, where one was subcutaneously administered phosphate buffer solution (PBS; control group, n=6∼7), while the other was subcutaneously administeredhUCB-MSCs (hUCB-MSC group, n=10∼11). The rats were allowed free access to food and water in a temperature- and humidity-controlled room (20℃, 60%) with a 12/12 h day/night cycle (7 am/7 pm). The rats were sacrificed using the carbon dioxide method [22]. All experiments followed the Guidelines on Ethical Standards for Investigation of Experimental Pain in Animals [23]. Additionally, the study protocol was approved by the Institutional Animal Care and Use Committee of Seoul National University. The official permission number for those animal experiments of the Institutional Animal Care and Use Committee of Seoul National University is SNU-081002-5.
Cell culture and transplantation
Human UCB-MSCs were obtained from umbilical cord blood immediately after delivery with consent from the mother. Cell culture of hUCB-MSCs was performed as previously described [12]. All procedures were approved by the Institutional Review Board of Seoul National University (IRB No. 1109/001-006). Two weeks after each neuropathic pain surgery, 1×105 hUCB-MSCs suspended in 50 µl PBS were injected subcutaneously into the plantar region of the left hind paw of each rat, using a 27G needle syringe connected to a Hamilton micro syringe for effective delivery of the cells. Control groups for each model were identically administered PBS.
Behavior tests
The rats were allowed to acclimatize to the test environment for at least 3 days before all behavior tests were conducted. We performed the von Frey test to assess changes in mechanical threshold after peripheral nerve injury and the effect of the hUCB-MSC treatment on pain-related behaviors. The rats were habituated to transparent plexiglass boxes (15×30×15 cm) placed on an elevated metal mesh platform for 20∼30 min. A series of ten von Frey monofilaments were applied to the entire plantar surface of the hind paw of CCI rats, the 3rd and 4th interdigit of SNL rats, and the external site of lateral plantar surface of the hind paw of SNI rats, which are innervated by the sural nerve [424]. The tests determined pain behavior such as paw withdrawal or licking response to three of five repetitive stimuli.
Immunohistochemistry
The animals were deeply anesthetized with 3% isoflurane via a nose cone approximately 2 to 3 min, and then, perfused transcardially with 200 to 300 ml saline, followed by 400 to 500 ml cold 4% paraformaldehyde in 0.1 M phosphate buffer (pH 7.4). The left DRG connected to L4 and L5, the L4∼L5 spinal cord, and the brain were dissected, and immunohistochemistry was performed as previously described [252627]. Positive cells area (%) presents the mean percentages of c-fos or CGRP or p-ERK or p-p38 or MMP-9 or MMP-2 or TIMP-2-immunoreactive (IR) neurons relative to the total number of neurons in the laminae I∼II layers of the ipsilateral L4∼L5 spinal dorsal horns.
Western blot
The animals were deeply anesthetized with 3% isoflurane via a nose cone for approximately 2 to 3 min, after which L4∼L5 spinal cord was quickly removed and immediately frozen in liquid nitrogen and stored at −70℃ until tissue lysis. The extracted proteins were heated at 95℃ for 5 min to denature the 3D structure of the protein and separated using 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis. With the help of an I-Blot transfer machine (Invitrogen, Waltham, MA, USA), the proteins were transferred to PVDF membranes and treated for 7 min. The membranes were blocked with 5% non-fat dry milk in Tris-buffered saline with Tween (TBST) and incubated overnight with primary antibody in TBST buffer at 4℃. The primary antibodies used were against MMP-2 (1:100, Santa Cruz Biotech, Dallas, TX, USA), MMP-9 (1:100, Santa Cruz Biotech, Dallas, TX, USA), TIMP-2 (1:100, Santa Cruz Biotech, Dallas, TX, USA) and alpha-tubulin (1:10000, Santa Cruz Biotech, TX, USA). The next day, the membranes were washed with TBST buffer several times and incubated with secondary antibody for an hour at room temperature. The horseradish peroxidase activity was measured using an enhanced chemiluminescence kit (ECL kit, Pierce, Thermoscientific, Rockford, IL, USA). The chemiluminescent signals were captured on autoradiography film.
Statistical analysis
The data are expressed as mean±standard error of mean (S.E.M.). SPSS version 17.0 software (SPSS Korea, Seoul, Korea) was used to compare the control and experimental groups. Statistical significance of difference was determined by Duncan's t-test. A p<0.05 was regarded as statistically significant.
Go to :
RESULTS
hUCB-MSCs decreased mechanical threshold in all three neuropathic pain models, with long-lasting antinociception in SNI model
Compared to the baseline value, mechanical threshold was decreased maximally 14 days after peripheral nerve injury in all neuropathic pain models (Fig. 1). We injected hUCB-MSCs at that time of maximally induced neuropathic pain. Mechanical threshold was significantly increased at 4 weeks after transplantation in both, CCI (Fig. 1A, p=0.00006) and SNL (Fig. 1B, p=0.032) models, and at 5 weeks after transplantation in SNI model (p=0.014). Furthermore, we observed threshold differences at 8 weeks, the longest observation time in this study, after transplantation in SNI model (Fig. 1C, p=0.0002).
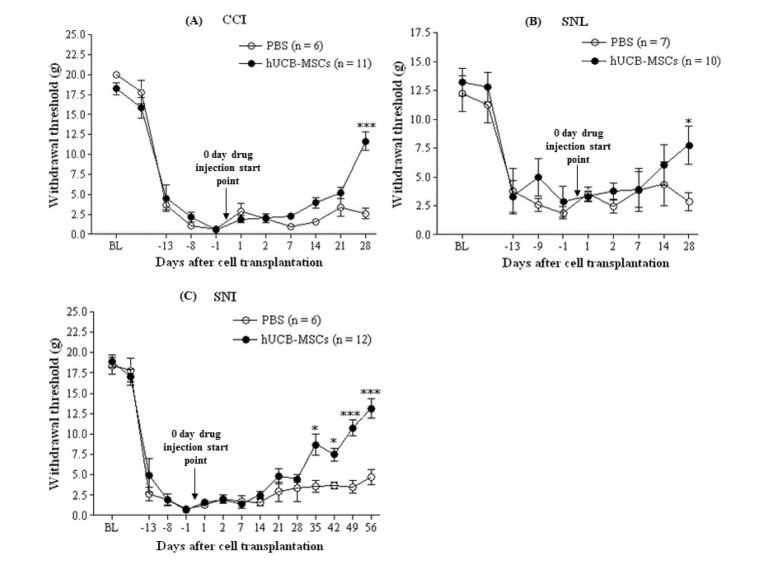 | Fig. 1The effect of hUCB-MSCs on mechanical hypersensitivity in the three neuropathic pain models.After establishment of neuropathic pain behavior (14 days after CCI, SNL, and SNI), PBS (n=6∼7) or hUCB-MSCs (n=10∼12, 1×105 cells/50 µl in PBS) were subcutaneously injected into the plantar. “0 day drug injection start point” of arrows stands for transplantation of hUCB-MSCs. Mechanical hypersensitivity measured by withdrawal threshold was assessed after 1 and 2 days, and every week for a month thereafter. In the three neuropathic pain models, mechanical threshold of the hUCB-MSCs group was higher than control group. BL indicates a withdrawal threshold at base line, which is the start point of neuropathic pain animal model establishment. The data are presented as mean±S.E.M. *p<0.05, ***p<0.001 indicate a significant difference compared to control group.
|
Transplanted hUCB-MSCs affected expression levels of pain-related molecules in the spinal cord determined via immunohistochemistry
As shown in Fig. 2, expression of c-fos decreased in the hUCB-MSCs grafts; however, this was significant only in CCI model (Fig. 2A, p=0.028 for CCI, p=0.059 for SNL, p=0.073 for SNI). CGRP expression decreased significantly in all models compared to in control animals (Fig. 2B, p=0.03 for CCI, p=0.003 for SNL, p=0.04 for SNI). Phosphorylated ERK expression decreased in the hUCB-MSCs-treated rats, but this was not statistically significant in SNI model (Fig. 2C, p =0.04 in CCI, p=0.0003 in SNL, p=0.1001 in SNI). p-p38 expression in the hUCB-MSCs-treated rats showed a significant decrease in CCI model (p=0.04) while p-p38 expression in the hUCB-MSCs-treated rats was not significant in SNL and SNI models (Fig. 2D, p=0.56 for SNL, p=0.78 for SNI).
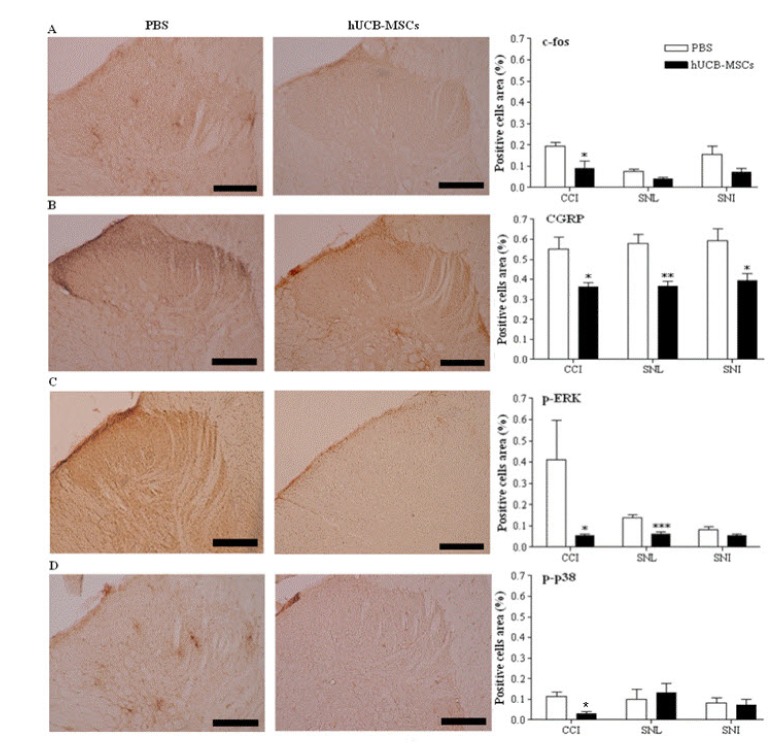 | Fig. 2Immunohistochemical analysis using the pain markers in the laminae I-II layers of the ipsilateral L4∼L5 spinal dorsal horns in the three neuropathic pain models.The laminae I∼II layers of the ipsilateral L4∼L5 spinal dorsal horns in CCI and SNL models were dissected and sampled on 4 weeks, while those layers in SNI models dissected on 8 weeks after transplantation of hUCB-MSCs. The number of cells positive for pain-related antibodies such as c-fos, CGRP, p-ERK and p-p38 was decreased in the hUCB-MSCs group (n=11 in CCI, n=10 in SNL, n=12 in SNI) compared to control group (n=6 in CCI, n=7 in SNL, n=6 in SNI). Positive cells area (%) presents the mean percentages of c-fos or CGRP or p-ERK or p-p38-immunoreactive (IR) neurons relative to the total number of neurons in the laminae I∼II layers of the ipsilateral L4∼L5 spinal dorsal horns. Statistical significance of difference was determined by Duncan's t-test. A p<0.05 was regarded as statistically significant. The data are presented as mean±S.E.M. *p<0.05, **p<0.01 and ***p<0.001 indicate a significant difference compared to control group. All of the micro-graphics are from the CCI group. A, c-fos; B, CGRP; C, p-ERK; D, p-p38. Scale bars, 200 µm
|
Transplanted hUCB-MSCs affected expression levels of MMP family proteins and TIMP-2 in the spinal cord, investigated by immunohistochemistry and western blot
MMP-9-positive cells were reduced in the hUCB-MSCs-treated rats compared to the control in CCI and SNI models, where significant difference was only observed in CCI model (Fig. 3A, p=0.008 for CCI, p=0.49 for SNI). MMP-9 expression was not significantly different in the hUCB-MSCs-treated rats in SNL model (p=0.83) too. MMP-2-positive cells were decreasedsignificantly in all three models compared to the control (Fig. 3B, p=0.049 for CCI, p=0.003 for SNL, p=0.015 for SNI). In contrast, TIMP-2-positive cells were increased in both, SNL and SNI models compared to control, but not in CCI model (Fig. 3C, p=0.397 for CCI, p=0.041 for SNL, p=0.02 for SNI). Western blotting revealed that hUCB-MSC transplantation induced TIMP2 protein expression in SNL and SNI models and reduced MMP2 protein expression in all three models (Fig. 3D). Although the results from western blotting differed between models, they were consistent with immunohistochemistry results.
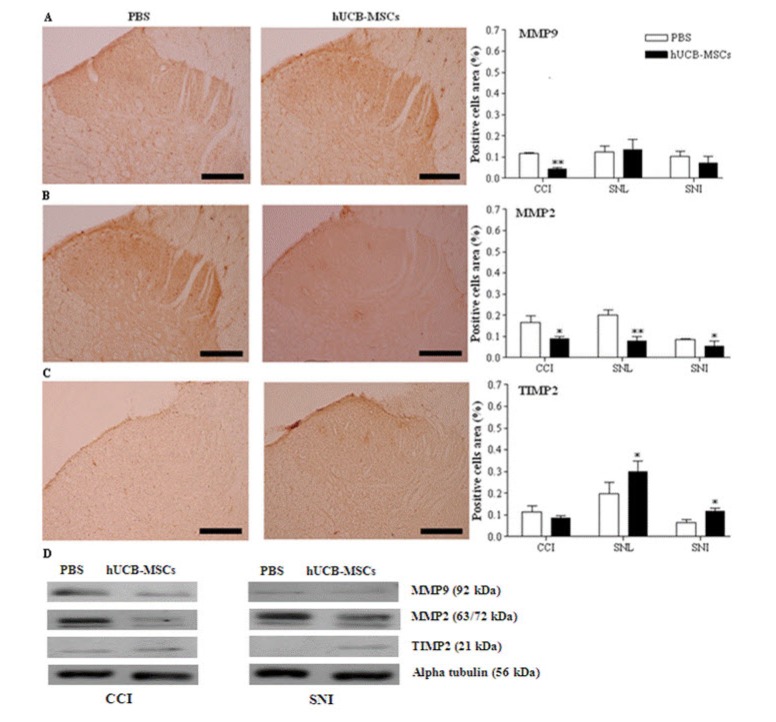 | Fig. 3Immunohistochemistry and western blot analysis for MMP-9, MMP-2, and TIMP-2 in the spinal cord.The laminae I-II layers of the ipsilateral L4-L5 spinal dorsal horns and L4-L5 spinal cord in CCI and SNL models were dissected and sampled on 4 weeks, while those layers and that spinal cord in SNI models dissected on 8 weeks after transplantation of hUCB-MSCs. The cells positive for MMP-9, MMP-2, and TIMP-2 in the laminae I-II layers of the ipsilateral L4-L5 spinal dorsal horn were analyzed in the three neuropathic pain models. Positive cells area (%) presents the mean percentages of MMP-9 or MMP-2 or TIMP-2 immunoreactive (IR) neurons relative to the total number of neurons in the laminae I-II layers of the ipsilateral L4-L5 spinal dorsal horns. Expression of MMP-9 was only reduced in CCI model. In all of the neuropathic pain models, expression of MMP-2 was significantly decreased in the hUCB-MSCs group (n=11 in CCI, n=10 in SNL, n=12 in SNI) compared to control group (n=6 in CCI, n=7 in SNL, n=6 in SNI). In the hUCB-MSCs group, TIMP-2-positive cells were significantly increased in SNL and SNI models. Results for expression of those antibodies was similar in the western blot analysis. Statistical significance of difference was determined by Duncan's t-test. A p<0.05 was regarded as statistically significant. The data are presented as mean±S.E.M. *p<0.05, **p<0.01 indicate a significant difference compared to control group. The micrographs A and B are taken from the CCI group and C from the SNL group. A: MMP-9, B: MMP-2, C: TIMP-2, D: western blot analysis. Scale bars, 200 µm.
|
Go to :
DISCUSSION
In this study, we showed that a single transplantation of hUCB-MSCs reduced mechanical allodynia 1 month after treatment, with a significant difference in peak levels in all three neuropathic pain models studied. This increased mechanical threshold in vivo was confirmed in terms of antibody expression levels.
Allodynia, a neuropathic pain symptom, is defined as pain evoked by normally innocuous stimuli, caused by reduced nociceptive thresholds, and is regarded more important than other clinical phenomena [2829]. The sensitization of neurons, which may occur within the dorsal horn, is characterized by increased spontaneous activity of dorsal horn neurons, increased responsivity to afferent input, and decreased threshold. To date,various pharmacological agents have been used to attenuate neuropathic pain, including N-methyl-D-aspartate receptor antagonists, gamma-aminobutyric acid agonists, opioid agonists, anticonvulsants[30]. However, clinical trials with these agents show limited efficacy, with risk of drug intolerance or addiction. Therefore, a new approach for neuropathic pain therapy is necessary and cell transplantation has emerged as a potential alternative. Previous studies have been reported regarding cell therapy in neuropathic pain models [31], primarily involving transplantation into the subarachnoid space [32333435]. Compared to stem cells of embryonic, bone marrow, and adipose origin, hUCB-MSCs have shown higher promise for clinical application. The merits of hUCB-MSCs include a higher number of mesenchymal progenitor cells per unit volume and lower carcinogenic risk [91011], as well as higher pluripotency and easy, painless acquisition [11]. Subcutaneous transplantation of hUCB-MSCs into the plantar region is preferred over other delivery methods such intravenous, intrathecal, or intracisternal injection due to ease of clinical application.
The present study was carried out in principal, rodent models of peripheral neuropathic pain. Using three different models, we circumvented the limitations of each and simultaneously achieved a comparison of their effects on hUCB-MSCs.
The transplanted hUCB-MSCs contributed to reduce pain-related behaviors such as paw licking, flinching, and shaking. Moreover, the in vivo data was supported by antibody expression patterns at the molecular level. ERK plays an important role in pain signal transduction and is extensively distributed in the central and peripheral nervous systems. Phosphorylated ERK expression is known to be activated sequentially in neurons, microglia, and astrocytes, which contributes to mechanical allodynia in an animal model for neuropathic pain [36]. Also, expression of p-ERK is increased by mechanical stimulation and noxious heat at the spinal cord dorsal horn level [3637]. Phosphorylation of ERK controls transcriptional regulation [38]. Subsequently, expression of c-fos and CGRP, markers of neuronal activation after noxious stimulation [39] is altered by p-ERK. Interestingly, ERK and p-38 are examples of mitogen-activated protein kinases; however, they differentially contribute to development of neuropathic pain in the spinal cord after peripheral nerve injury [40]. For example, phosphorylation of p38 in the spinal cord is essential for early phase of neuropathic pain[41], whereas phosphorylation of ERK is important for both early and late phase of neuropathic pain [36]. Our immunohistochemical analysis results are in agreement with these previous reports, wherein expression of c-fos, CGRP, and p-ERK decreased slightly in hUCB-MSCs grafts with p-p38 expression not being affected. We hypothesized that these phenomena were due to trophic factors secreted by hUCB-MSCs, especially MMPs and TIMPs [1721]. Another previous study reported that early and late phases of neuropathic pain are associated with different proteins from the MMP family. In nerve injury-induced neuropathic pain models in rats and mice, MMP-9 shows a transient upregulation in injured DRG primary sensory neurons and spinal microglia during early-phase neuropathic pain, whereas MMP-2 shows a delayed response in DRG satellite cells and spinal astrocytes during late-phase [2140]. Our results showed that MMP-2 expression decreased, with TIMP-2 expression simultaneously increasing in the transplanted group. This is in accordance with other studies showing that MMP-2 activity is regulated by its endogenous inhibitor, TIMP-2 [4243]. Thus, TIMP-2 was secreted by the hUCB-MSCs–inhibited activation of MMP-2, and this may have attenuated the neuropathic pain condition. Moreover, previous studies have shown that transplanted hUCB-MSCs improve motor function or pain-related behaviors in the rodent spinal cord injury model [1044], which is probably contributed by the effect of MMP-2 and TIMP-2 [45].
The LC is essential in pain modulation [46]. It has bilateral projections, which descend primarily to the spinal dorsal horn laminae I∼II and V, where antinociceptive effect can be observed [47]. Electrical or chemical stimulation of the LC inhibits responses to noxious stimuli in the spinal dorsal horn neurons via release of noradrenaline. Thus, induction of antinociception was confirmed based on previous findings regarding c-fos expression [4748]. Moreover, it is correlated with behavioral recovery.
We demonstrated that hUCB-MSCs showed a significant improvement in animal models for neuropathic pain after intraplantar, subcutaneous transplantation. It seems that hUCB-MSCs transplantation cause secretion of TIMP-2, which inhibits MMP 2 activation that otherwise produces neuropathic pain symptoms, via IL-βcleavage and activation of p-ERK in astrocytes [21]. This finding was indirectly confirmed by expression of c-fos and CGRP, which are generally used as stress markers p-ERK, which is upstream of c-fos and CGRP. Among three animal models for neuropathic pain, spinal cord cells positive for c-fos, CGRP, p-ERK, p-p 38, MMP-9 and MMP 2 were significantly decreased in only CCI model of hUCB-MSCs-grafted rats. The CCI model has been extensively used for many neuropathic studies because it closely mimics the clinical nerve injury conditions and pain nature such as complex regional pain syndrome type 2 [245].
In conclusion, our current study shows that transplantation ofhUCB-MSCs is a novel strategy to relieve neuropathic pain, with potential as an alternative clinical therapy. However, further studies are required to determine clinical feasibility for long-term use to rule out adverse effects due to immune response.
Go to :
 XML Download
XML Download