

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Atractylodis Rhizoma (Cangzhu) is widely used as a traditional herbal medicine [12]. Cangzhu contains essential oils, phenolic acids, sesquiterpenes, and polyethylene alkynes [345]. Atractylodin, one of the polyethylene alkynes with Hepatoprotective, anti-oxidative, and anti-obesity effects, is used to ameliorate rheumatic diseases and night blindness [67]. Atractylodin is also used to improve digestive disorders and the delay of L-NNA-induced gastric emptying [8].
Inflammatory bowel disease (IBD) and irritable bowel syndrome (IBS) are major intestinal disorders and both IBD and IBS are characterized by having inflammation and the co-occurring intestinal dysmotility in different extent [910]. Diarrhea, constipation and intestinal inflammation are the main symptoms of the patients with IBD and IBS [1112]. Clinical and epidemiological studies indicate that the inflammation is found from the jejunum to the rectum in both IBS and IBD [1314151617]. Diarrhea is the common symptoms of IBD; IBS may have constipation-predominant symptom or diarrhea-predominant symptom [181920]. The differential diagnosis for distinguishing IBD and IBS based on clinical signs and symptoms are difficult [212223]. The above observations suggest that amelioration of intestinal inflammation and co-occurring dysmotility are required for the treatment of IBD and IBS. However, simultaneous improvement of intestinal inflammation and the co-occurring dysmotility using current available drug in clinic is rarely reported.
Our pre-experiments indicated that atractylodin ameliorated intestinal inflammation and exerted beneficial modulation on intestinal contractility in rats. The present study was designed to characterize the effects of atractylodin on intestinal inflammation and the co-occurring intestinal dysmotility and to reveal the mechanisms by using both diarrhea prominent (DP) rat and constipation prominent (CP) rat models.
Go to : 
METHODS
Animals
Forty male rats of Sprague–Dawley (SD) weighing 180~220 g were provided by the Experimental Animal Center of Dalian Medical University (Dalian, China). The experiment protocol was carried out based on the Declaration of Helsinki, and supported by Dalian Medical University Animal Care and Ethics Committee. Every six rats were housed in a cage and put in a temperature-controlled room with a 12-hlight-dark cycle. Food and water were available for ad libitum consumption.
Experimental models of diarrhea and constipation
DP rats were established by intracolonic instillation of 4.0% (V/V) acetic acid and restraint stress, and the control rats underwent by intracolonic instillation with saline; CP rats were established by daily gavage with cool water (0℃~4℃) for 14 days and the control rats gavaged with water at room temperature [2425262728]. The successful establishment of both DP and CP rats was confirmed by the determination of intestinal inflammation in both rat models and by the observation of the increased jejunal contractility in DP rats and decreased jejunal contractility in CP rats. Intragastric gavage atractylodin (10.0 mg/kg) for 7 days were performed for both DP and CP rats and normal rat controls were given same amount of vehicle. Jejunal segments isolated from control rats, DP rats, and CP rats were collected and used in the determination of jejunal contractility and evaluate the effects of atractylodin [29]; the jejunal segments isolated from DP rats, atractylodin-treated DP rats, CP rats, atractylodin-treated CP rats, and control rats were used in the determination of mRNA expression of myosin light chain kinase (MLCK), protein content of MLCK, and phosphorylation extent of the 20 kDa myosin light chain (p-MLC20). The granules and moisture content of the feces from each group were calculated daily.
ELISA assay
Serum pro-inflammatory cytokines and mediators, including tumor necrosis factor-alpha (TNF-α), Interleukin-1-beta (IL-1β), and Interleukin-6 (IL-6) were determined using double-antibody sandwich ELISAs (R&D Systems, USA).
Intestinal histopathology
The segments of jejunum were detached to assess the morphological changes of the jejunum in CP and DP rats. The collected segments of jejunum were fixed and stained with hematoxylin and eosin (H&E).
Immunofluorescence determination
The jejunal tissue slices were deparaffinized and rehydrated by using xylene and graded concentrations of ethanol. The slices were incubated with 3% H2O2 (10 min) and permeabilized with 0.3% Triton X-100 in PBS for 15 min. The slices were then blocked with 10% goat serum for 45 min at 37℃, followed by incubation with primary rabbit anti-NF-κB antibody or rabbit anti-iNOS antibody overnight at 4℃. After rinsing, the slices were covered with goat anti-rabbit secondary antibodies conjugated with a fluorophore for 1 h at 37℃. After rinsing in PBS, the slices were covered with 1 µg/ml DAPI for 7 min and then determined using fluorescence microscope.
Tissue preparation and contractility determination
Jejunal segments were isolated from the intact jejunum of normal, CP, and DP rats respectively as described previously [30]. Jejunum was cut into approximately 2.0 cm in length. One end of jejunal segment was fixed to the wall of the tissue bath chamber (20.0 mL volume), and the other end was connected to a force-displacement transducer in longitudinal direction. Contractile amplitude of isolated jejunal segments was recorded and identical time-interval of each assay with same start and stop time was chosen to compare the amplitude of contractions before and after drug treatment in all assays [31]. Jejunal segments isolated from CP and DP rats and assayed in normal Krebs buffer were selected as a pair of low-high contractile states. The contractility of the normal jejunal segment measured in normal Krebs buffer was chosen as the normal contractile state.
The other five pairs of low/high jejunal contractile states were established by pre-incubation in modified Krebs buffer containing adrenaline (5.0 µM)/acetylcholine (ACh) (5.0 µM) or low Ca2+ (1.3 mM)/high Ca2+ (5.0 mM) or low K+ (2.5 mM)/high K+ (10.0 mM) or low Na+ (100.0 mM)/high Na+ (150.0 mM) or nitric oxide donor sodium nitroprusside (SNP) (5.0 µM)/erythromycin (10.0 µM).
Western blot analysis
The contents of myosin regulatory light chain phosphorylation (p-MLC20) and myosin light chain kinase (MLCK) in the jejunal segment were examined by using Western blot analysis [3233]. The jejunal segments were frozen and stored in liquid nitrogen for determination. The ground product was incubated for 30 min in ice-cold homogenization buffer. The blots on nitrocellulose filter membrane were probed with phosphor-myosin light chain 2 (Ser 19) antibody (1:1000) and myosin light chain 2 (total myosin light chain) antibody (1:1000) respectively at 4℃ with gentle shaking, overnight. For MLCK determination, the blots on nitrocellulose filter membrane were probed with MLCK antibody (1:1000). Anti-rabbit IgG secondary antibodies were used at 1:1000 for 90 min at room temperature and bands were detected and quantified by using a Multi Spectral Imaging System (UVP, Cambridge, UK).
Real-time PCR analysis of MLCK expression
Total RNA was extracted from the jejunal segment by using TRIzol® reagent (Life Technologies, Carlsbad, CA, USA). Total RNA detach and Real-time PCR for mRNA quantization were performed as previously described [34]. For cDNA synthesis, oligo dT primers were used to prime the RT reactions. The primers (Takara Biotechnology (Dalian) CO., LTD., China) were designed to determine the transcripts of MLCK according to published sequences. The house-keeping gene glyceraldehyde-3-phosphate dehydrogenase (GAPDH) served as an internal control. The primers sequences for PCR were as follows: MLCK: 5′-AAT GGT GTT GCT GGA GAT CGA GGT-3′ (forward) and 5′-GCT GGA TCA AAT TGC GGT GGT TCA-3′ (reverse), GAPDH : 5′-GGC AAG TTC AAT GGC ACA GT-3′ (forward) and 5′-TGG TGA AGA CGC CAG TAG ACT C- 3′ (reverse). RT - PCR was performed using SYBR green fluorescence during amplification on an ABI 7500 Fast Real-Time PCR System (Applied Biosystems, Carlsbad, CA, USA). PCR conditions were as follows: initial denaturation at 95℃ for 30 s, 40 thermal cycles (95℃ for 5 s, 61℃ for 34 s, and 72℃ for 30 s). The mRNA levels of MLCK were normalized to the mRNA levels of the housekeeping gene GAPDH. The comparative Ct method (2−ΔΔCt) was used to analyze the differences of MLCK mRNA expression between each group [35].
Reagents
Atractylodin with purity of 98% was obtained from Chengdu Mustbio Technology co., Ltd., China. MLCK antibody and GAPDH antibody were purchased from Abcam (Cambridge, UK). Tumor necrosis factor-alpha (TNF-α), Interleukin-1-beta (IL-1β), and Interleukin-6 (IL-6) double-antibody sandwich ELISAs were purchased from Sigma (St. Louis, MO, USA). Anti-rabbit IgG secondary antibody was bought from Cell Signaling Technology (CST, Beverly, MA, USA). 4,6-diamidino-2-phenylindole (DAPI) was provided by Sigma (St. Louis, MO, USA).
Statistical analysis
The results are shown as mean±standard deviation (SD). Statistical analysis was performed using unpaired Student's test in the organ chamber experiment, and with one-way ANOVA, followed by Bonferroni's post hoc test in the other experiments. p<0.01 indicates a statistically significant difference between groups.
Go to :
RESULTS
The establishment of CP and DP rat models
CP and DP rat models were used in preclinical studies [273637]. In the present study, whether CP and DP models were successfully established was confirmed by analysis of the following parameters: granules/moisture contents of feces, jejunal contractility, and jejunal inflammation.
In day 7, the granules and moisture contents of feces from CP and DP rats were significantly decreased and increased respectively, compared with normal controls, indicating the successful establishment of CP and DP rat models. Atractylodin significantly increased the decreased granules and moisture contents of feces of CP rats and significantly decreased those increased parameters of DP rats (Table 1).
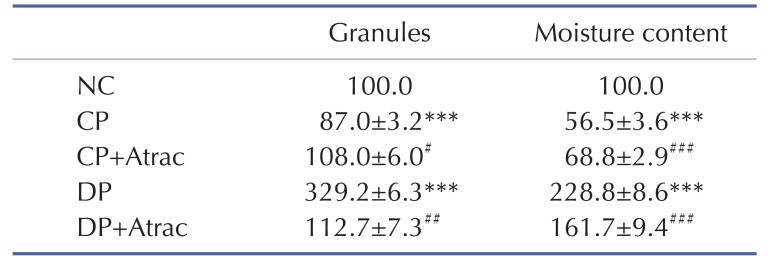
Jejunal inflammation was found in both CP and DP rats; decreased jejunal contractility was observed in CP rats and increased jejunal contractility was observed in DP rats (Fig. 1).
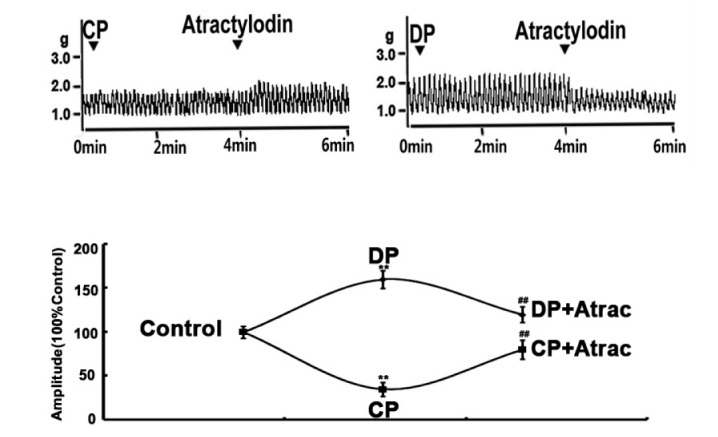 | Fig. 1Atractylodin-exerted contractile-state-dependent regulation on rat jejunal contractility.Representative traces of atractylodin-exerted stimulatory and inhibitory effects on rat jejunal contractility in CP and DP rats, respectively. Contractile amplitude of jejunal segments in normal contractile state is set to a relative value of 100% (control). Other data are the relative values compared with control. **p<0.01 compared with the NC; ##p<0.01 compared with contractile amplitude of jejunal segments in DP group or CP group before being given atractylodin.
|
Anti-inflammatory effects of atractylodin on CP and DP rats
Neutrophil infiltration and proinflammatory cytokines levels may lead to inflammation. In the present study, neutrophil infiltration in H&E-stained jejunal sections and proinflammatory cytokines levels were determined. As is shown in Fig. 2, neutrophil infiltration and proinflammatory cytokines TNF-α, IL-1β, and IL-6 contents were significantly increased in both CP and DP rats, compared with the controls. Atractylodin treatment significantly reversed the jejunal inflammation-related parameters in both CP and DP rats.
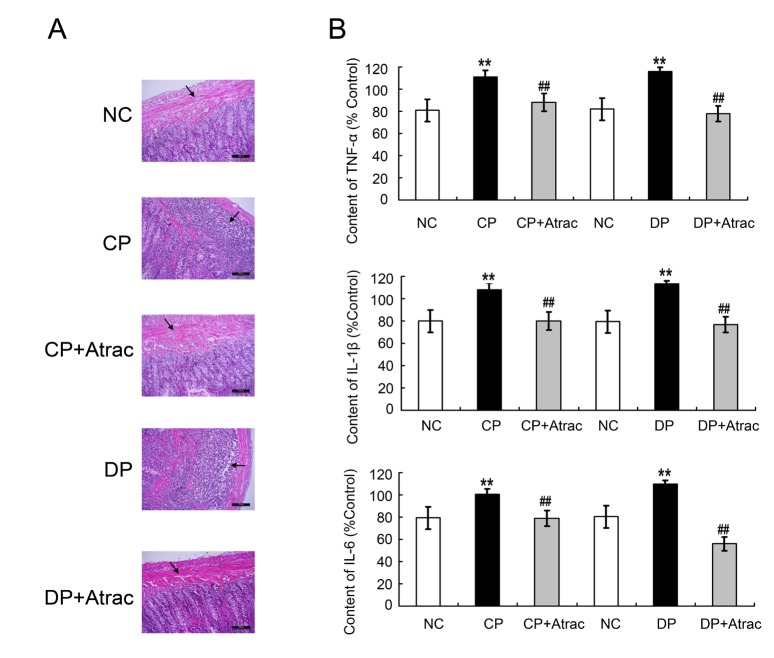 | Fig. 2Histomorphological characteristics of atractylodin-induced anti-inflammatory effects on DP and CP rats.Panel A: the histology images including normal jejunal tissue, CP jejunal tissue, DP jejunal tissue, CP+Atrac jejunal tissue, and DP+Atrac jejunal tissue. Panel B: statistical analysis of the effects of atractylodin on the serum contents of TNF-α, IL-1β and IL-6 in both DP and CP rats. Data are expressed as mean±SD (% NC, n=6); **p<0.01 compared with the normal controls; ##p<0.01 compared with CP group or DP group before being given Atractylodin, respectively. Atrac, atractylodin; NC, normal control rats; CP, constipation prominent rats; DP, diarrhea prominent rats.
|
NF-κB is involved in inflammatory processes and the expression of the inducible nitric oxide synthase (iNOS) is one of the direct consequences of inflammation [3839]. The results obtained from immunofluorescence determination indicated that the treatment with atractylodin significantly reduced the expression of NF-κB and iNOS in jejunal segments in both DP and CP rats compared with untreated CP and DP controls (Fig. 3), suggesting that atractylodin can ameliorate intestinal inflammation in both CP and DP rats.
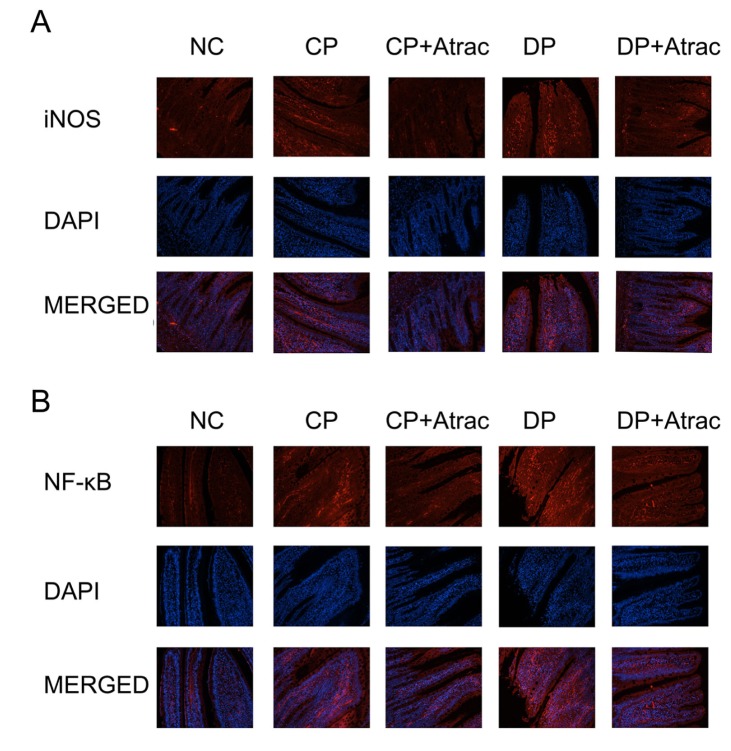 | Fig. 3Effect of atractylodin on iNOS and NF-κB in CP and DP rats.Panel A: histology images of iNOS expression (red fluorescence), DAPI nuclear staining (blue fluorescence), and merged images of the immunofluorescence of iNOS and DAPI. Panel B: histology images of NF-κB expression, DAPI nuclear staining, and merged images of the immunofluorescence of NF-κB and DAPI. DAPI, 4′,6-diamidino-2-phenylindole. Cells viewed at ×200 magnification.
|
Atractylodin-exerted contractile-state-dependent regulation
Based on our pre-experiment, 5.0 µM atractylodin was the chosen as the optimized concentration in characterizing the effects of atractylodin on the contractility of jejunal segments in both CP and DP rats. The jejunal contractility of CP and DP rats was decreased and increased respectively. Atractylodin (5.0 µM) enhanced the decreased jejunal contractility of CP rats and reduced the increased jejunal contractility of DP rats (Fig. 1).
To verify whether atractylodin exerted regulation on jejunal contractility is contractile-state dependent, 5 pairs of low/high contractile states were established to make the evaluation. Jejunal contractility in the 5 low contractile states induced by adrenaline, low Ca2+ or low Na+ or low K+ or SNP was significantly decreased and jejunal contractility in the 5 high contractile states induced by ACh or high Ca2+ or high Na+ or high K+ or erythromycin was significantly increased, compared with the normal jejunal controls. Atractylodin significantly enhanced the jejunal contractility in all five low contractile states respectively and significantly decreased the jejunal contractility in all five high contractile states respectively (Fig. 4), suggesting that atractylodin-exerted regulation is characterized by a contractile state dependent manner.
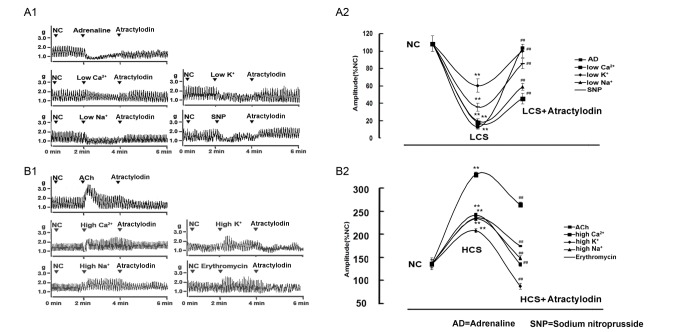 | Fig. 4The characteristic of atractylodin-exerted contractile state dependent contractile-state-dependent regulation.Panel A: Representative traces (A1) and statistical analysis (A2) of atractylodin-induced stimulatory effects on the contractility of jejunal segment in 5 low contractile states. Panel B: Representative traces (B1) and statistical analysis (B2) of atractylodin-induced inhibitory effects on the contractility of jejunal segment in 5 high contractile states. Contractile amplitude of jejunal segments in normal contractile state is set to a relative value of 100% ( normal control, NC). Other data are the relative values compared with NC. Data are expressed as mean±SD (% NC, n=6); **p<0.01 compared with the NC; ##p<0.01 compared with contractile amplitude of jejunal segments in LCS (low contractile state) or HCS (high contractile state) before being given atractylodin, respectively. NC, normal control; SNP, sodium nitroprusside; AD, adrenaline; ACh, acetylcholine; ERY, erythromycin.
|
Role of myosin phosphorylation in atractylodin-exerted regulation
Phosphorylation of 20 kDa regulatory myosin light chain (p-MLC20) plays a key role in initiating the contraction of smooth muscle. p-MLC20 extent, MLCK protein content, and mRNA expression of MLCK were significantly decreased and increased in CP and DP rats respectively in comparison with those parameters in their controls. Atractylodin significantly up-regulated the decreased p-MLC20 extent, MLCK protein content, and mRNA expression of MLCK in jejunal segments in CP rats and down-regulated those increased parameters in DP rats (Fig. 5).
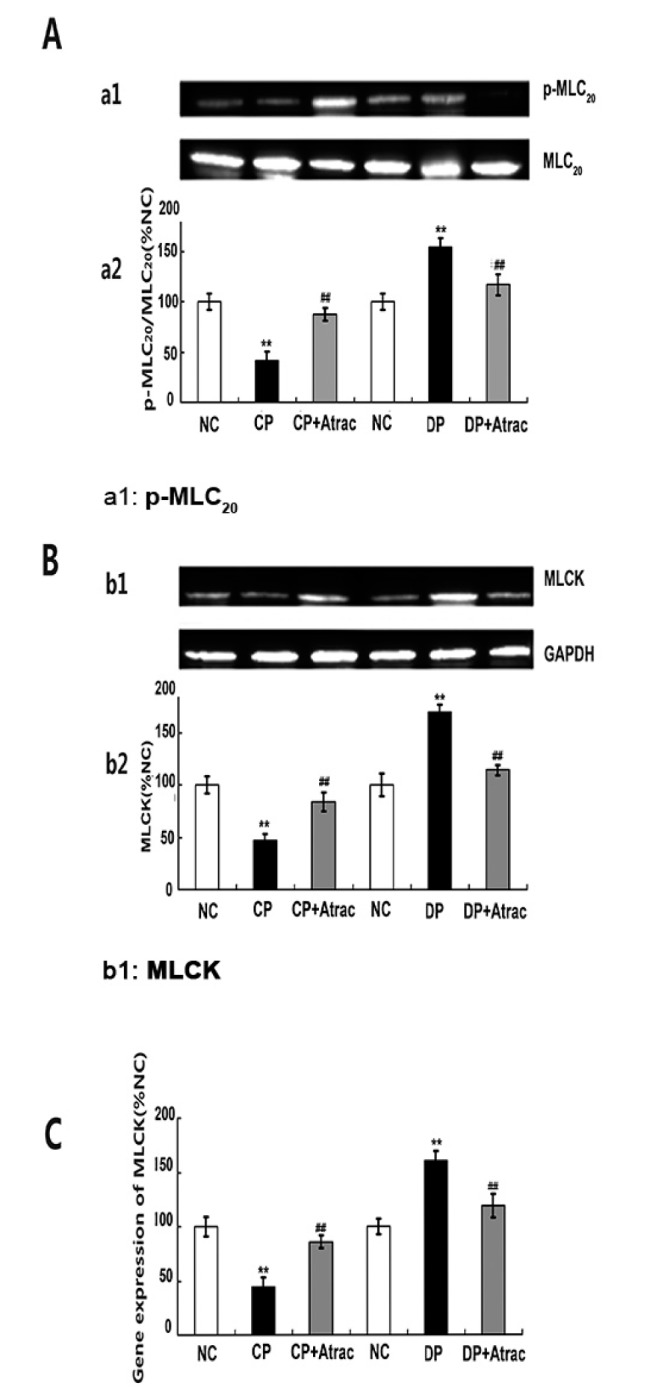 | Fig. 5Effects of atractylodin on myosin phosphorylation-related mechanisms.Panel A: phosphorylation extent (a1) and statistical analysis (a2) of 20-kDa myosin light chain (p-MLC20), Panel B: protein contents (b1) and statistical analysis (b2) of myosin light chain kinase (MLCK), Panel C: MLCK mRNA expression in jejunal segment in normal control (NC) rats, atractylodin-treated CP rats (CP+Atrac), and atractylodin-treated DP rats (DP+Atrac). Data were expressed as the mean±SD from four independent experiments; **p<0.01 compared with the normal controls; ##p<0.01 compared with CP group or DP group before being given atractylodin, respectively. p-MLC20, phosphorylated 20-kDa myosin light chain; MLC20, 20-kDa myosin light chain; GAPDH, glyceraldehyde-3-phosphate dehydrogenase.
|
Go to :
DISCUSSION
Based on the clinical evidence that both IBD and IBS have intestinal inflammation and dysmotility in different extent, the differential diagnosis for distinguishing IBD and IBS based on clinical signs and symptoms are difficult, and approximately 60~80% of IBD patients suffer from IBS [40], how to ameliorate both intestinal inflammation and co-occurring dysmotility simultaneously are the tough challenges in the clinical practice. However, none of the current available medications including anti-inflammatory drugs, immunosuppressive drugs, 5-aminosalicylic acid, corticosteroids, and antibiotics used for the treatment of IBD and IBS is found to simultaneously improve intestinal inflammation and co-occurring dysmotility (including both increased and decreased contractility). To cope with the challenge, the present study was designed to establish both CP and DP rat models with co-occurring intestinal inflammation in both models to investigate the effects of atractylodin on the jejunal inflammation and co-occurring intestinal dysmotility.
Firstly, the evidence of jejunal inflammation and decreased jejunal contractility in CP rats, and jejunal inflammation and increased jejunal contractility in DP rats indicated the successful establishment of CP and DP rat models. In accordance with the observation that inflammation is found in small and large intestines in both IBS and IBD [2941], the inflammation was also found in small and large intestines in both CP and DP rats in our study. As the atractylodin-exerted contractile-state-dependent regulation on jejunal segments was not observed on rat colon segments, jejunal segments were used to evaluate atractylodin-exerted regulation on inf lammation and co-occurring dysmotility. The physiological significance of atractylodin-exerted different effects on jejunum and colon needs further study.
Secondly, inflammation is reported to play a major role in the pathogenesis of gastrointestinal disorders [42]. Intestinal mucosal inflammation induced intestinal dysmotility and impaired the barrier function [4344]. Furthermore, the inflammatory mediator, NF-κB, regulates the expression of genes involved in inflammation and plays a key role in both first pro-inflammatory phase and later regulation of the resolution of inflammation [4546]. Proinflammatory cytokines (TNF over-expression of TNF–α, IL-1β, and IL-6 contribute to IBD [4748]. Inhibition of the expression of NF-κB, TNF-α, and pro-inflammatory mediator iNOS, ameliorates IBD [46]. Our study indicated that atractylodin ameliorated jejunal epithelial inflammation by inhibiting pro-inflammatory cytokines TNF-α, IL-1β, and IL-6 and inflammatory mediators iNOS and NF-κB in both CP and DP rats, suggesting its potential clinical implication for amelioration of the intestinal inflammation in both IBD and IBS.
Thirdly, atractylodin (one-membered ring) exerted stimulatory effect on the jejunal segments isolated from CP rats (low contractile state) and exerted inhibitory effect on jejunal segments isolated from DP rats (high contractile state) respectively, showing the characteristics of contractile-state-dependent regulation rather than structure-activity relationship. Atractylodin-induced contractile-state-dependent regulation is not an exception. Our previous studies indicated that ginsenoside Re (four-membered ring), emodin (three-membered ring), and diprophylline (two-membered ring) were also found respectively to have contractile-state-dependent regulation on rat jejunal contractility [495051].
To further characterize atractylodin-induced contractile-state-dependent regulation, 5 pairs of low/high contractile states were established in the study. Atractylodin significantly enhanced the jejunal contractility in all 5 low contractile states. Atractylodin significantly reduced the jejunal contractility in all 5 high contractile states. These results indicated that atractylodin-induced regulation is characterized by contractile-state-dependent manner regardless the cause which induces low or high contractile state.
MLCK is essential for gastrointestinal motility [52]. MLCK phosphorylates 20 kDa regulatory myosin light chain (MLC20) and p-MLC20 interact with actin, inducing the contraction of smooth muscles [53]. Atractylodin up-regulated the decreased p-MLC20, MLCK contents and mRNA expression of MLCK in the jejunal segments isolated from CP rats and down-regulated the those parameters in the jejunal segment isolated from DP rats, suggesting atractyldin-induced contractile-state-dependent regulation is positively correlated to p-MLC20 and MLCK expression.
To summarize, atractylodin simultaneously improved jejunal inflammation and co-occurring dysmotility in both CP and DP rats characterized by reducing the over-expression of inflammatory cytokines and increasing and decreasing the contractility and p-MLC20 in jejunal segments isolated form CP and DP rats respectively, suggesting that atractylodin may have potential application for improving intestinal inflammation and the co-occurring dysmotility.
Go to :
 XML Download
XML Download