

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Inflammation is a complex immunobiological response used to fight off pathogens such as bacteria, viruses, and fungi. These pathological stimuli induce various processes, including stimulating immune cells to migrate to the injured tissues and to produce inflammatory mediators and cytokines under activation conditions [12]. One of the important molecular events that regulates such responses is the activation of nuclear factor (NF)-κB [23]. Thus, when inflammation occurs upon recognition of surface receptors by pathogen-derived molecules such as lipopolysaccharide (LPS), cytokines, or sterile inflammation-inducing molecules (including high-mobility group box 1 protein (HMGB1)), many intracellular proteins in immune cells are activated by subsequent phosphorylation to deliver the signals from outside the nucleus into the nucleus via activation and translocation of transcription factors such as NF-κB [4]. NF-κB is involved in the mRNA expression of numerous inflammatory genes, such as inducible NO synthase (iNOS), cyclooxygenase (COX)-2, and tumor necrosis factor (TNF)-α [5]. However, if the inflammation does not cease, the sustained response causes serious damage to the body. Indeed, many previous studies have shown that chronic inflammation is a major contributor to various serious diseases, such as cancer, diabetes, atherosclerosis, and autoimmune diseases [6]. Therefore, effective suppression of chronically processed inflammation is regarded as an important strategy to prevent various diseases.
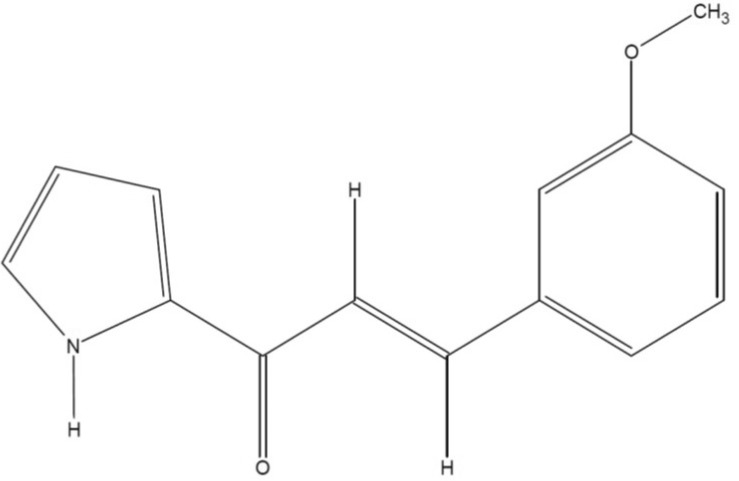
Several propenone structures (e.g., 1-furan-2-yl-3-pyridin-2-yl-propenone) are reported to have anti-inflammatory effects in various inflammatory responses [78]. Recently, our group designed a series of compounds with a propenone structure. One of these compounds, (E)-3-(3-methoxyphenyl)-1-(2-pyrrolyl)-2-propenone (MPP, Fig. 1), is a derivative synthesized from (E)-1-aryl-3-(2-pyrrolyl)-2-propenones by aldol condensation. Although a structurally similar class of propenone-type compounds are anti-inflammatory, and there are plenty of derivatization sites in this compound, the biological activity of MPP has not yet been elucidated. Here, we report the anti-inflammatory activity of MPP using macrophage-derived inflammatory responses.
Go to : 
METHODS
Materials
Polyethylenimine (PEI), 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (a tetrazole) (MTT), NG-nitro-L-arginine methyl ester L-NAME, PP2, piceatannol (Picea), and lipopolysaccharide (LPS, E. coli 0111:B4) were purchased from Sigma Chemical Co. (St. Louis, MO, USA). Fetal bovine serum (FBS) and RPMI1640 were obtained from GIBCO (Grand Island, NY, USA). Dulbecco's modified Eagle's medium (DMEM) was obtained from Thermo Fisher Scientific Inc. (Waltham, MA, USA). RAW264.7 and HEK293 cells were purchased from ATCC (Rockville, MD, USA). All other chemicals used in this study were of analytical grade from Sigma Chemical Company. Phospho-specific or total-protein antibodies recognizing p65, p50, inhibitor of κBα (IκBα), AKT, Src, spleen tyrosine kinase (Syk), lamin A/C and β-actin were obtained from Cell Signaling Technology (Beverly, MA, USA).
Preparation of (E)-3-(3-methoxyphenyl)-1-(2-pyrrolyl)-2-propenone
MPP was prepared, according to previously [9]. Briefly, 2-Acetylpyrrole (0.83 g, 7.6 mmole) and 3-methoxybenzaldehyde (1.03 g, 7.6 mmole) were added to an ice-cold solution of 2 M-NaOH (5 ml) and ethanol (7 ml). The resulting solution was gradually brought to room temperature and stirred for 5 h, followed by heating at 40℃ for 30 min. The solution was neutralized with 1 M-HCl aqueous solution to pH 8~9 and then the resulting precipitate was collected and recrystallized from ethanol. The yield of the purified product was 83% and mp was 144℃.
Expression vectors and DNA transfection
DNA constructs encoding Src, Syk, MyD88, TRIF, AP-1-Luc, and NF-κB-Luc were used as reported previously [1011]. The sequences of all constructs were confirmed by automated DNA sequencing. Expression vectors were transfected in HEK293 cells by the PEI method in 12-well plates (1×106 cells/ml), as reported previously [1213]. After 24 h, the transfected cells were treated with MPP for an additional 24 h before termination.
Cell culture and drug preparation
RAW264.7 cells (passage No.: 10 to 20) were cultured in RPMI 1640 with 10% heat-inactivated FBS and 1% penicillin/streptomycin at 37℃ in 5% CO2. HEK293 cells were maintained in DMEM media supplemented with 5% heat-inactivated FBS and 1% penicillin/streptomycin at 37℃ in 5% CO2. The stock solutions of MPP for the in vitro experiments were prepared in dimethylsulfoxide (DMSO).
Determination of NO production
After pre-incubation of RAW264.7 cells (1×106 cells/ml) for 18 h, MPP was added to the cells for 30 min. The cells were then treated with LPS (1 µg/ml) for 24 h. The inhibitory effects of MPP (0 to 50 µM) or L-NAME (0.5 to 1.5 mM) on the production of NO were determined by analyzing NO levels using Griess reagents [1415].
Cell viability test
The cytotoxic effects of MPP (0 to 50 µM) were evaluated using a conventional MTT assay, as previously reported [161718]. For the final 3 h of culture, 10 µl of MTT solution (10 mg/ml in phosphate-buffered saline, pH 7.4) were added to each well. Reactions were stopped by the addition of 15% SDS into each well to solubilize the formazan. The absorbance at 570 nm (OD570-630) was measured using a Spectramax 250 microplate reader (BioTex, Bad Friedrichshall, Germany).
mRNA analysis through semiquantitative reverse transcriptase (RT)-polymerase chain reaction (PCR)
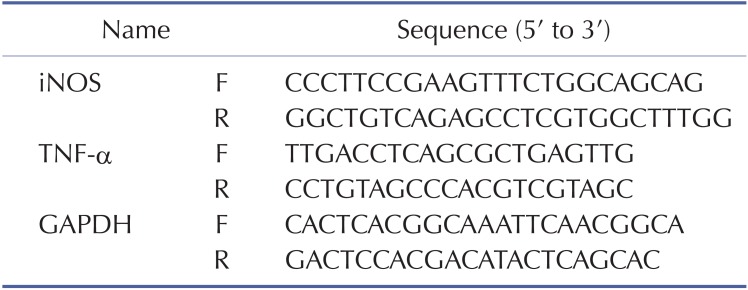
To evaluate cytokine mRNA expression levels, RAW264.7 cells pre-treated with MPP (0~50 µM) for 30 min were incubated with LPS (1 µg/ml) for 6 h. Total RNA was isolated using TRIzol Reagent (Gibco BRL) according to the manufacturer's instructions, and stored at –70℃ until further use. Semiquantitative RT reactions were conducted as previously reported [19]. The primers (Bioneer, Seoul, Korea) used in these experiments are listed in Table 1 (F: forward, R: reverse).
Luciferase reporter gene activity assay
HEK293 cells (1×106 cells/ml) were transfected with 1 µg of plasmid containing NF-κB-Luc or AP-1-Luc along with β-galactosidase using the PEI method in a 24-well plate, according to the manufacturer's protocol. The cells were used for experiments 24 h after transfection. Luciferase assays were performed using the Luciferase Assay System (Promega), as reported previously [20].
Preparation of cell lysates and nuclear fractions for immunoblotting
RAW264.7 or HEK293 cells (5×106 cells/ml) were washed three times in cold PBS and lysed in a lysis buffer, as reported previously [21]. Nuclear lysates were prepared using a three-step procedure [22]. After treatment, the cells were collected with a rubber policeman, washed with 1×PBS, and lysed in 500 µl of the lysis buffer on ice for 4 min. During the second step, the pellet (the nuclear fraction) was washed once with wash buffer without Nonidet P-40. During the final step, the nuclei were resuspended in an extraction buffer consisting of the lysis buffer plus 500 mM KCl and 10% glycerol. The nuclei/extraction buffer mixture was frozen at –80℃ then thawed on ice and centrifuged at 14,000 rpm for 5 min. The supernatant was collected as the nuclear extract. Whole-cell or nuclear lysates were then analyzed by a conventional immunoblotting method [23]. The total and phosphorylated levels of p65, p50, IκBα, AKT, Src, Syk, HA, Myc, lamin A/C, and β-actin were visualized using an ECL system (Amersham, Little Chalfont, Buckinghamshire, UK), as reported previously [24].
Statistical analyses
All of the data presented in this paper are expressed as the mean±SD. For statistical comparisons, results were analyzed using either the ANOVA/Scheffe's post-hoc test or the Kruskal-Wallis/Mann-Whitney test. A p-value less than 0.05 was considered to be statistically significant. All statistical tests were carried out using the computer program, SPSS (SPSS Inc., Chicago, IL, USA).
Go to :
RESULTS
Effects of MPP on NO production and cell viability in LPS-treated RAW264.7 cells
In order to confirm the effects of MPP as an anti-inflammatory drug, its capacity to reduce NO production under LPS-treated conditions was analyzed using macrophage-like RAW264.7 cells. The production of NO was dose-dependently inhibited by MPP (0 to 50 µM) in LPS-treated RAW 264.7 cells (Fig. 2A). Meanwhile, L-NAME, an iNOS inhibitor, displayed an effective suppressive pattern under the same NO production conditions, implying that our conditions are well established. In addition, no cytotoxic activity of MPP at its effective anti-inflammatory dose was observed, indicating that the inhibition of NO production by MPP is not due to non-specific activity (Fig. 2B).
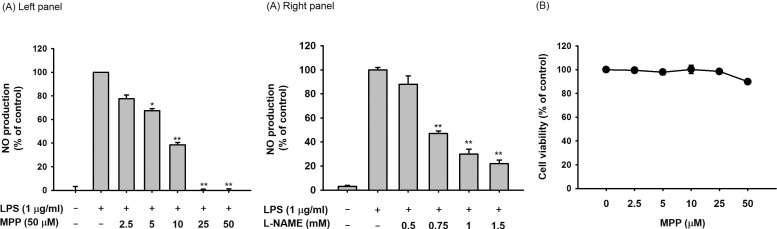 | Fig. 2Effect of MPP on the production of NO from LPS-treated RAW 264.7 cells and on the viability of RAW264.7 cells.(A) RAW264.7 cells (1×106 cells/ml) were stimulated with LPS (1 µg/ml) in the presence or absence of MPP (0 to 50 mM) or L-NAME (0 to 1.5 µM) for 24 h. The supernatants were then obtained, and the NO concentration in the supernatants was confirmed using the Griess assay. (B) RAW264.7 cells (1×106 cells/ml) were treated with MPP for 24 h, and cell viability was examined using the MTT assay. All data are expressed as the mean±SD of experiments, which were performed with six samples. *p<0.05 and **p<0.01 compared to normal or control groups.
|
Effects of MPP on transcriptional activation
To determine whether the NO suppressive activity of MPP occurs at the transcriptional level, the mRNA expression levels of inflammatory genes such as iNOS and TNF-α, which are known to be mediated by NF-κB, were assayed using semiquantitative RT-PCR. As seen in Fig. 3A, the mRNA levels of iNOS and TNF-α were repressed by MPP in LPS-treated RAW264.7 cells in a dose-dependent manner. To further confirm the effects of MPP on transcriptional activation, a luciferase reporter gene activity assay was employed. HEK293 cells were co-transfected with NF-κB-Luc reporter genes under the overexpression of the adaptor molecules TRIF and MyD88, as reported previously [2526]. Under these conditions, cells exhibited increased levels of luciferase activity up to 140- and 900-fold in the presence of TRIF and MyD88, respectively (Figs. 3B and 3C). Interestingly, MPP dose-dependently suppressed the upregulated luciferase activity when used at 25 and 50 µM (Figs. 3B and 3C). Similarly, nuclear translocation of p65, a major submit of NF-κB, was also reduced by MPP treatment (25 µM) for 30 min (Fig. 3D), implying that the blockade of p65 translocation could be linked to the suppression of NF-κB-mediated transcription of inflammatory genes.
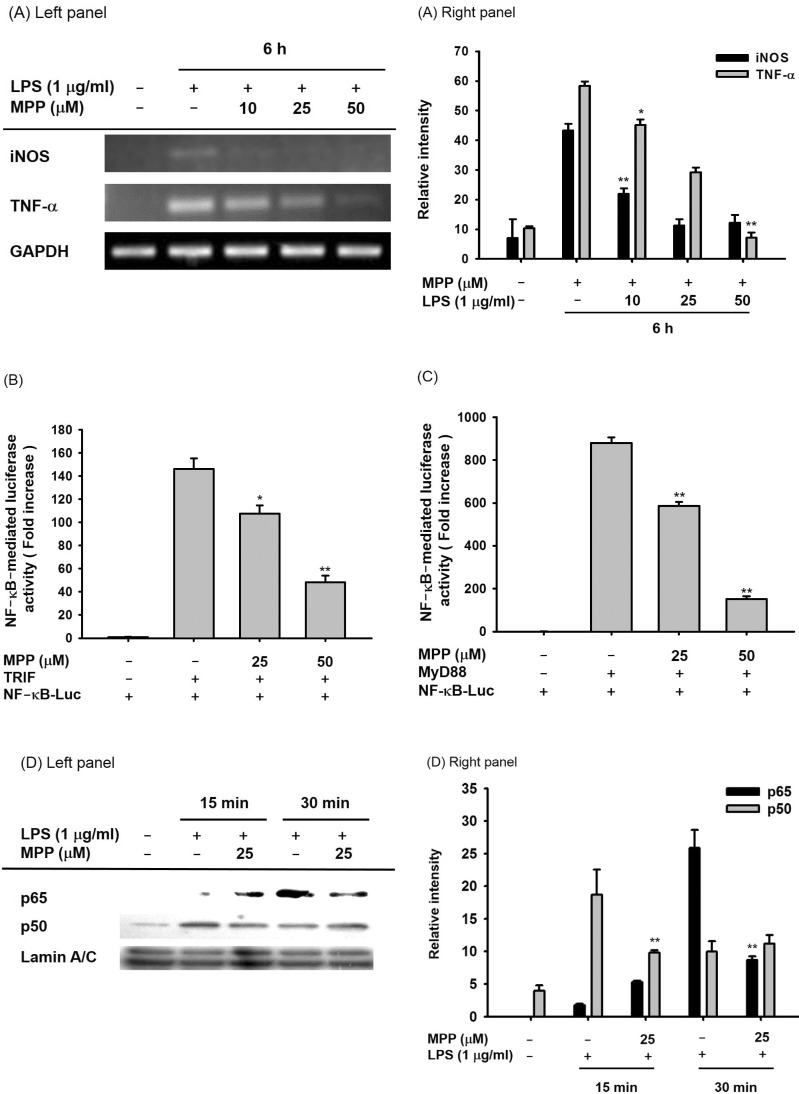 | Fig. 3Effects of MPP on transcriptional activation in LPS-stimulated RAW 264.7 cells and MyD88/TRIF-treated HEK293 cells.(A) The mRNA levels of iNOS and TNF-α in LPS (1 µg/ml)-treated RAW264.7 cells in the presence or absence of MPP (10, 25 and 50 µM) were determined by RT-PCR. (B and C) HEK293 cells co-transfected with NF-κB-Luc plasmid construct and β-gals in the presence or absence of MyD88 or TRIF were treated with MPP (25 and 50 µM). Luciferase activity was measured using a luminometer. (D) The nuclear levels of p65 and p50 in LPS (1 µg/ml)-stimulated RAW264.7 cells in the presence or absence of MPP (25 µM) were detected by immunoblotting analysis. Relative intensity was calculated using total levels by the DNR Bio-imaging system. Data (B and C) are expressed as the mean±SD of experiments, which were performed with six samples. *p<0.05 and **p<0.01 compared to normal or control groups.
|
Effects of MPP on the upstream signaling cascade for NF-κB activation
It is known that the translocation of NF-κB is regulated by complicated intracellular signaling cascades composed of Src, Syk, AKT and IκBα [23]. In order to determine the MPP targeted signaling molecules, the levels of the phospho- and total forms of these molecules were evaluated by immunoblotting assay. The phospho-forms of IκBα and AKT in MPP-treated cells were reduced at 30 min (Fig. 4A). Subsequently, MPP inhibited the phospho-protein levels of Syk and Src, which are upstream signaling enzymes that activate AKT (Fig. 4B).
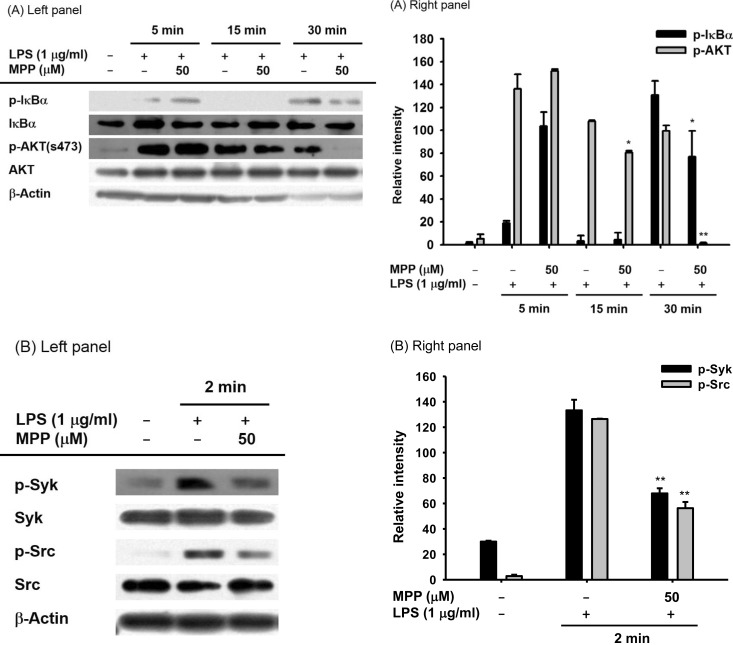 | Fig. 4Effects of MPP on the upstream signaling cascade for NF-kB activation in LPS-stimulated RAW 264.7 cells.(A and B) RAW264.7 cells were incubated with LPS (1 µg/ml) in the presence or absence of MPP (50 µM) for 30 min. After preparation of whole cell lysates, the total and phospho-protein levels of Src, Syk, AKT, and IκBα from the lysates were detected by immunoblotting analysis. Relative intensity was calculated using total levels by the DNR Bio-imaging system. *p<0.05 and **p<0.01 compared to normal or control groups.
|
Effects of MPP on autophosphorylation of overexpressed Src and Syk
Since increased levels of Src and Syk phosphorylation are a result of the autophosphorylation of their kinase domains [27], we next validated whether MPP could suppress the autophosphorylation of Src and Syk in cells overexpressing these genes. For this, HA-Src and Myc-Syk were transfected in HEK293 cells and increased autophosphorylation levels were observed (Fig. 5), as reported previously [28]. As we expected, MPP (25 and 50 µM) also exhibited suppressive activity on the autophosphorylation of these proteins (Figs. 5A and 5B). Meanwhile, the significant roles of Src and Syk in inflammatory responses were confirmed by treating cells with specific inhibitors of these kinases (PP2 and piceatannol, respectively). Thus, the production of NO in LPS-treated RAW264.7 cells was clearly reduced by PP2 and piceatannol, implying that these enzymes play important roles in NO production pathway.
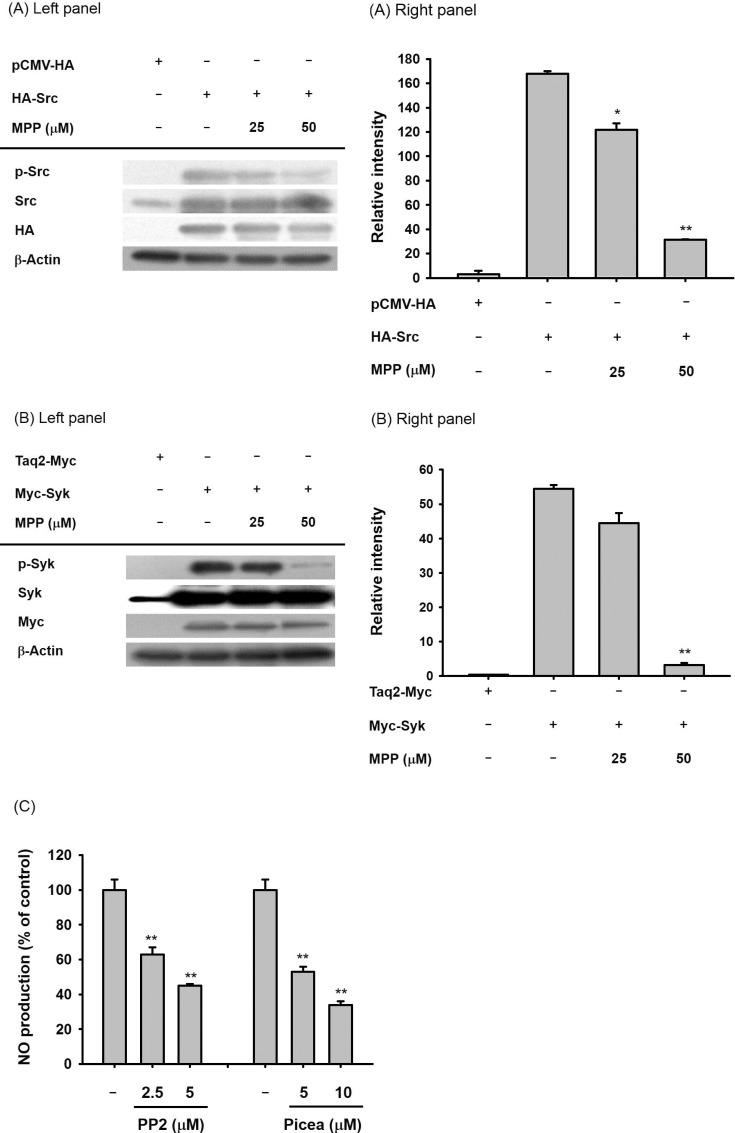 | Fig. 5Effects of MPP on autophosphorylation of Src and Syk in Src- or Syk-transfected HEK293 cells.(A and B) Inhibitory activity of MPP (25 and 50 µM) on the autophosphorylation of Src or Syk in Src- or Syk-transfected HEK293 cells was measured by immunoblotting analysis. (C) RAW264.7 cells (1×106 cells/ml) were stimulated with LPS (1 µg/ml) in the presence or absence of PP2 (2.5 and 5 µM) or piceatannol (Picea, 5 and 10 µM) for 24 h. The supernatants were then obtained, and the NO concentration in the supernatants was confirmed using the Griess assay. Relative intensity was calculated using total levels by the DNR Bio-imaging system. *p<0.05 and **p<0.01 compared to normal or control groups.
|
Go to :
DISCUSSION
Our results strongly imply that MMP has significant anti-inflammatory properties. NO production in RAW 264.7 cells decreased in a dose-dependent manner without affecting cell viability upon MMP treatment (Figs. 2A left panel and 2B). Moreover, the mRNA expression levels of the pro-inflammatory genes iNOS and TNF-α were reduced (Fig. 3A) in MPP-treated cells. In addition, the fact that MPP has significant suppressive effects on the NF-κB signaling pathway, as evidenced by results of the luciferase reporter gene assays (Figs. 3B and 3C) and immunoblotting analysis with nuclear fractions (Fig. 3D), strongly suggests that NF-κB is an important target transcription factor in MPP-mediated anti-inflammatory activity. It is widely known that NF-κB is one of the major transcription factors regulating inflammatory responses. Consequently, higher levels of NF-κB have identified in numerous inflammatory diseases, including colitis, hepatitis, gastritis, and arthritis [2930]. Inhibition of this transcription factor with plant extracts such as Cinnamomum cassia, Archidendron clypearia, and Evodia lepta, as well as with naturally-occurring and synthetic chemicals such as Scutellarein, 21-O-angeloyltheasapogenol E3, and (5-hydroxy-4-oxo-4H-pyran-2-yl)methyl 6-hydroxynaphthalene-2-carboxylate, has been linked to the amelioration of inflammatory diseases, including gastritis, colitis, hepatitis, and pancreatitis [252831323334]. Therefore, it has been speculated that the NF-κB-inhibitory activity of MPP could critically contribute to its immunopharmacological action in inflammatory responses.
The down-regulation of nuclear p65/NF-κB translocation by MPP (Fig. 3D) strongly supports the idea that MPP could affect the upstream inflammatory process of NF-κB activation in the cytoplasm. In fact, NF-κB translocation requires the phosphorylation of IκBα by IKK which allows for the degradation of this protein [35]. The phosphorylation of IκBα is also mediated by a series of upstream signaling cascades composed of Akt, PDK1, PI3K, Src, and Syk during the activation of toll like receptors (TLRs) [2336]. According to our results (Fig. 3D), MPP was found to suppress the nuclear levels of p65, implying that MPP could target some of the signaling enzymes involved in IκBα phosphorylation and subsequent p65 translocation. Indeed, the fact that the inhibition of phosphorylation of Akt by MPP occurred 30 min post-MPP addition (Fig. 4A) and that the phosphorylation of Src and Syk was also downregulated by MPP at 2 min implies that NF-κB is not directly suppressed by MPP, but rather that this compound might suppress upstream enzymes involved in NF-κB activation [37]. Interestingly, several points strongly indicate that Src and Syk could be target proteins in MPP-mediated anti-inflammatory action. Src and Syk are the major NF-κB regulatory protein tyrosine kinases [3839], and are involved in the regulation of IκBα phosphorylation [40] through AKT regulation at early time points (within 5 min of stimulation) as we have seen. Transfection of the Syk and Src genes was able to increase the phosphorylation levels of Syk and Src. Indeed, upregulated levels of autophosphorylated Src and Syk upon overexpression from Myc-Syk or HA-Src in HEK293 cells were clearly observed and, interestingly, MPP was found to suppress these events (Figs. 5A and 5B). Moreover, PP2 and piceatannol displayed strong inhibitory action on NO production under the same conditions (Fig. 5C), implying that a blockade of Src and Syk by MPP or by their specific inhibitors (PP2 and piceatannol) could contribute to the downregulation of NF-κB-dependent inflammatory activity. In addition, we previously identified the phospho-forms of Src and Syk in the stomach, liver, and intestine under inflammatory conditions [4142], indicating that these enzymes could also participate in the induction of organ inflammatory symptoms. Therefore, these data along with previous findings strongly suggest that MPP-mediated anti-inflammatory activity could be managed by the inhibition of Src and Syk linked to NF-κB activation. Since it is important to know whether MPP is able to directly suppress the enzymatic activity of Src and Syk, we will measure this possibility using a direct enzyme assay in the future.
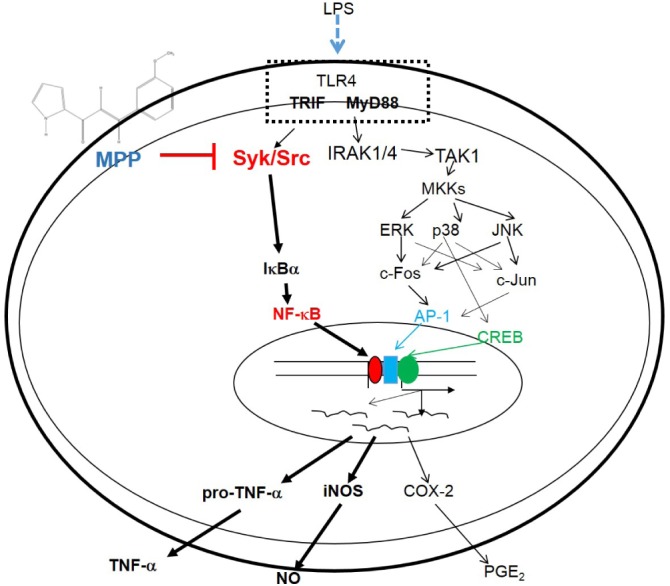
In summary, we found that MPP is capable of inhibiting NO production and inflammatory gene (iNOS and TNF-α) expression, and of diminishing a series of molecular inflammatory responses, including the phosphorylation of Src, Syk, Akt, and IκBα, and the translocation of p65/NF-κB, as summarized in Fig. 6. Because MPP displayed a strong anti-inflammatory effect, this compound should be considered for further development as an effective anti-inflammatory drug for treating various inflammatory symptoms. Therefore, relevant work will be pursued in future studies. In addition, to improve the potency of MPP, other derivatives of MPP will be continuously prepared and evaluated.
Go to :
 XML Download
XML Download