

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Synaptic plasticity, which is the strengthening or the weakening of the synaptic response to stimulus, has attracted considerable attention from neuroscientists investigating various aspects of brain function. Long-term potentiation (LTP) and long-term depression (LTD) are the two major forms of synaptic plasticity observed in electrophysiology and other investigative studies examining at the synapses of the neurons [1]. The hippocampus, especially the region of Schaffer collateral–CA1 projection fibers, is the most well-studied region in the central nervous system for synaptic plasticity, and the mechanisms of LTP and LTD have already been well established. Synaptic plasticity in the hippocampus is one of the most widely recognized cellular and synaptic model of learning and memory in the neuroscience field and continues to be adapted in different behavioral or disease models for various brain regions.
LTD is the activity-dependent reduction in the efficacy and strength of synaptic transmission lasting for a few hours or longer. There are two major forms of LTD in the CNS: the N-methyl-D-aspartate receptor (NMDAR)-dependent LTD [23] and NMDAR-independent or mGluR-dependent LTD [456]. These can be induced by different stimulating protocols in different regions and have been best investigated in the hippocampus and cerebellum.
mGluR-dependent LTD was first observed at the synapses of the cerebellar parallel fiber—Purkinje cells [789]. mGluR-dependent LTD involves the activation of mGluRs, which can be classified into three groups: Group I comprises mGluR1 and mGluR5, which are mainly present in the postsynaptic site [10] and have been mainly investigated in the context of LTD; Group II includes mGluR2 and mGluR 3; and Group III consists of mGluR4, mGluR6, mGluR7, and mGluR8. Groups II and III mGluRs are mainly expressed in the presynaptic site. Previous studies have shown that mGluR-dependent LTD can be induced after activation of mGluR1, mGluR5, mGluR2, mGluR3, and mGluR7 [6]. In this review, we will focus on mGluR-dependent LTD, especially in the cortical regions.
Go to : 
PROTOCOLS TO INDUCE mGluR LTD
Two major methods are used to induce mGluR-dependent LTD. First is the application of chemicals that stimulate mGluRs. Acute administration of Group I mGluR agonist 3,5-dihydroxyphenylglycine (DHPG) is the most common method to induce mGluR-dependent LTD throughout the brain [1112]. Group II mGluR needs to be activated in some regions and Group II mGluR agonist 2S, 2'R, 3'R)-2-(2', 3'-dicarboxycyclopropyl) glycine (DCG-IV) is one of the most well-known drugs capable of inducing LTD in the prefrontal cortex [1314].
The second method of inducing mGluR-dependent LTD is low-frequency stimulation (LFS). 900 single pulses in 1 Hz usually induces NMDAR-dependent LTD, and 900 pairs of 50-ms-apart pulses in 1 Hz are known to induce mGluR-dependent LTD [151617]. In the CA1 region of the hippocampus, paired-pulse LFS (PP-LFS) activates Group I mGluRs and M1 muscarinic acetylecholine receptors [151819]. However, in some cortical regions, such as the anterior cingulate cortex (ACC) and insular cortex (IC), 900 single pulses in 1 Hz induce LTD that are fully blocked by (S)-α-methyl-4-carboxyphenylglycine (MCPG), which is an antagonist to Groups I and II mGluRs [202122]. In addition, there are also other unique protocols such as the application of mGluR agonists and electrical stimulation with tetanus [2324] or using weaker LFS and maintaining the postsynaptic potential at –40 mV which is called the pairing protocol [2526].
Go to :
mGluR LTD MECHANISM
LTD dependent on Group I mGluR is the most studied form of mGluR-dependant LTD throughout the brain and has been extensively investigated by neuroscientists. Stimulation of Group I mGluR activates phospholipase C (PLC), inositol triphosphate (IP3) pathway to release Ca2+ from intracellular stores and protein kinase C (PKC) [272829]. PKCα phosphorylates ser880 of GluA2 to trigger endocytosis of α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor (AMPAR) and reduce the level of surface expression [29303132]. Several other protein elements such as p38 mitogen-activated protein kinase (p38 MAPK) [3334
35], extracellular signal-regulated kinases (ERKs) [36], Arc [3738], striatal-enriched protein phosphatase (STEP) [29394041], phosphoinositide 3-kinase (PI3K) [42], protein tyrosine phosphatases (PTP) [34] are also known to be involved in Group I mGluR-dependent LTDs. Several detailed reviews of the mechanism of mGluR-dependent LTD have been published previously [6124344].
Go to :
CORTICAL mGluR-DEPENDENT LTD
mGluR-dependent LTD in the hippocampus and the cerebellum have been extensively reviewed previously [6124344]. Here, we present an overview of the less-reviewed cortical regions.
Anterior Cingulate Cortex
The anterior cingulated cortex (ACC) is known to participate in a wide variety of functions in the brain such as cognition, error detection, decision making, memory, emotion, and pain [45464748
49]. Numerous human brain imaging studies have investigated the role of ACC in these functions, however, studies on synaptic plasticity in animals are mainly restricted to the pain field. The ACC consists of five layers (I, II, III, V, and VI) and the excitatory pyramidal neurons are mainly positioned in the layer II/III and V. Layer II/III pyramidal neurons mostly receive sensory inputs from the medial thalamus and project to layer V/VI neurons. Layer V pyramidal neurons receive inputs from layer II/III and the thalamus and send projections to other cortical regions. Numerous local interneurons such as parvalbumin (PV) and somatostatin (SOM) positive interneurons are localized in layer I and II-VI [5051]. Field recordings in rat and mice have shown that LTD is induced in the ACC with 1-Hz single-pulse LFS for 15 min and had similar results with several pharmacological studies [2022]. Field recordings in rat were performed in the layer II, and the recordings in the mouse was done throughout the ACC layers. Treatment with MGPC (500 µM), an antagonist to mGluR of Groups I and II, inhibited this LTD in both rat and mice, but the NMDAR antagonist APV (50 µM) was not able to completely block LTD. Unlike the case with the hippocampus, single-pulse LFS in the neural populations of the ACC induced strong synaptic depression, and the NMDAR function was less important. Moreover, the mGluR5 antagonist MPEP (10 µM) had no effect on the ACC neurons, whereas the mGluR1 antagonist LY 367385 (100 µM) fully blocked this LFS-induced LTD. L-type voltage gated calcium channel (L-VGCC) blocker also inhibited the LTD in the ACC induced by single-pulse LFS. These results imply that single-pulse LFS induced by LTD in the ACC depends on mGluR1 and L-VGCC (Fig. 1A). LTD was also induced by combined treatment with DHPG (100 µM) and MPEP (10 µM) for 20 min. When a digit of hind paw or 2.5 cm of the tail was amputated, LTD induced by LFS as well as chemical LTD induced by DHPG+MPEP were impaired. Chronic peripheral injury was involved in the disruption of the mGluR-dependent synaptic plasticity in the ACC, and this could be one of the functional characteristics in a phantom pain models brain. By priming the ACC slice of the amputated animal with low-dose DHPG (20 µM) and MPEP (10 µM) before the application of LFS, the impairment in the LTD was rescued [20]. This priming effect was PKC dependent. This type of metaplastic application could be a possible direction for the further development of treatment strategies in pain-related situations.
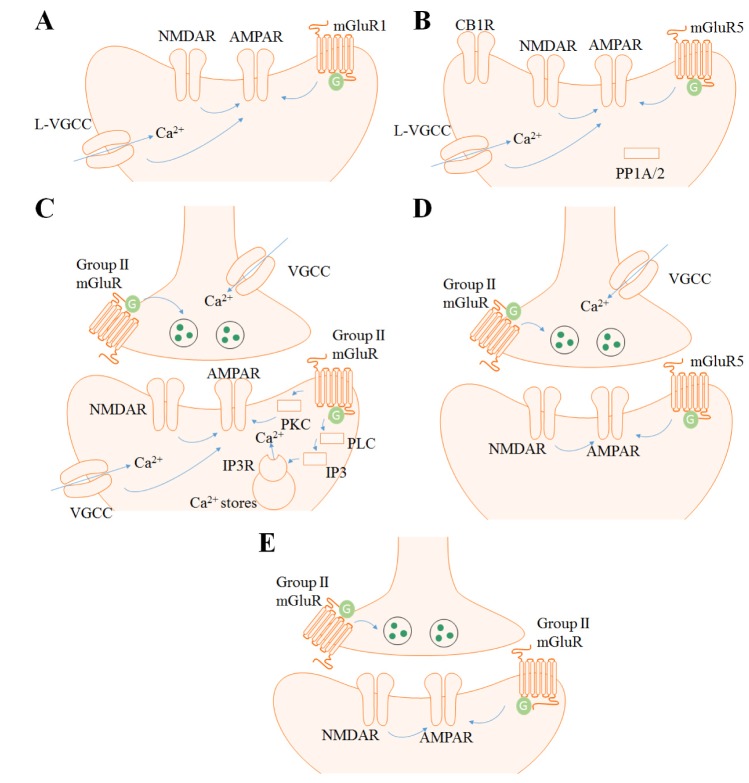 | Fig. 1Models of cortical mGluR-LTDs.(A) mGluR LTD in the ACC is mGluR1, L-VGCC and partially NMDAR dependent. (B) mGluR LTD in the IC is mGluR5, NMDAR, L-VGCC, CB1R and PP1/2A dependent. (C) mGluR LTD in the PFC is Group II mGluR, PLC, IP3, PKC and NMDAR dependent. (D) mGluR LTD in the perirhinal cortex is mGluR5, Group II mGluR and NMDAR dependent. (E) mGluR LTD in the visual cortex is Group II mGluR and NMDAR dependent.
|
Insular Cortex
IC is also a region known for its diverse functions, including perception, motor control, self-awareness, and cognitive functioning. Several behavioral studies such as those pertaining to conditioned taste aversion [525354] and pain [555657] have been conducted in the past. However, only few IC studies address synaptic plasticity [585960]. IC is divided into granular (GI), dysgranular (DI) and agranular (AI) cortices. All are composed of five layers as the ACC except the GI which has six layers including layer IV [61]. Pyramidal neurons are found mainly in the layer III and V [62] and there are also GABAergic interneurons in the layer V [63]. The afferent and efferent projections of the IC are well studied throughout the layers in several animal models although the cell type composition within layers still needs to be investigated further.
Two studies [2164] was performed in a manner similar to that of Kang et al. 2012. Recordings were done throughout all the layers in the IC and was plotted as superficial (layer I~II/III) and deep layer (layer V~VI). LTD in the IC was induced by 1-Hz single-pulse LFS for 15 min, and this LTD was blocked by APV (50 µM), nimodipine (10 µM), a selective cannabinoid 1 receptor (CB1R) antagonist AM251 (5 µM) and a selective protein phosphatase 1/2A (PP1/2A) inhibitor okadaic acid (1 µM). Interestingly, it was also fully blocked by MPEP (10 µM) but not CPCCOEt (100 µM), a selective mGluR1 antagonist. Therefore, LFS induced LTD in the IC is dependent on NMDAR, L-VGCC, CB1R, PP1/2A and mGluR5, when the same stimulating protocol induced mGluR1-dependent LTD in the ACC (Fig. 1B). DHPG (100 µM, 20 min) was also able to induce chemical LTD in the IC. This LTD was blocked when nimodipine (10 µM) was administered 20 min before DHPG infusion, thereby implying the importance of L-VGCC in DHPG-LTD. There was no layer difference in the experiments.
When the tail was amputated, LFS induced LTD was blocked in IC as ACC, but chemical LTD induced by DHPG remained unchanged [64]. Therefore, tail amputation might have more effect in the NMDAR than mGluR5 in IC. However, the impaired LFS-induced LTD could be improved by mGluR-dependent metaplasticity. Application of DHPG (20 µM) had no effect at the baseline, but subsequent LFS was able to induce LTD in the IC slice of the amputated mice. This priming effect was PKC dependent.
Prefrontal Cortex
The prefrontal cortex (PFC) is a region known for working memory [656667], attention [686970] and executive function [717273]. Numerous studies have investigated this region on several topics and is a popular research target because of its variety of functions. There are many more studies on mGluR-dependent LTD in PFC than in the ACC or IC; and few will be reviewed here. PFC consists of five layers as the ACC. Layer I contains neuropil, axons and GABAergic interneurons. Layers II/III and V/VI are composed of pyramidal neurons and various interneuron types [74].
Most of the mGluR LTD in the PFC is dependent on Group II mGluR. The Group I mGluR agonist DHPG (100 µM, 10~15 min) was not able to induce LTD in the prefrontal slice, but acute bath application of the potent Group II mGluR agonist DCG IV (50~100 nM) showed depression of the synaptic response for more than 40 min in layer I-II to layer V pyramidal neuron glutamatergic synapses of rat PFC [75]. This mGluR-dependent LTD was blocked when the postsynaptic cell was injected with Ca2+ chelator BAPTA (100 mM). It was also blocked when APV (100 µM) was applied together with DCG IV [14]. Moreover, the phospholipase C inhibitor U-73122 (4 µM, bath application), IP3 receptor blocker heparin (4 mg/ml in recording electrode targeting postsynaptic neuron), PKC inhibitor RO318220 (0.2 µM, bath application), all impaired the LTD. These results suggest that intracellular calcium, NMDAR-mediated responses, phospholipase C, IP3 receptor, and PKC together contribute to LTD induced by DCG IV (Fig. 1C).
LTD induced by DCG IV (0.2 µM, 10 min) in layer V pyramidal neurons was impaired when rats were treated with cocaine repeatedly for more than 5 days [13]. This impairment of LTD was improved when selective D1-like receptor antagonist SCH23390 (0.5 mg/kg) was coadministered with cocaine. Moreover, bilateral intra-mPFC infusion of PKC inhibitor bisindolylmaleimide I (0.4 nmol/side) or adenosine A3 receptor antagonist MRS1220 (0.5 nmol/side) also improved the impaired LTD. These results indicate that repetitive cocaine exposure inhibits Group II mGluR function via D1-like receptor, PKC, and A3 receptor.
A study on the importance of mGuR3 in LTD at PFC layer V pyramidal neurons and fear extinction has also been published [76]. Bath application of Group II agonist LY379268 (0.1 µM, 10 min) was able to induce LTD in the mice PFC slice. This LTD was blocked when mGluR3-negative allosteric modulator VU0469942 (10 µM) or VU0477950 (10 µM) was applied. When VU0477950 (3~100 mg/kg) was intraperitonially injected, fear extinction learning was impaired.
Perirhinal Cortex
The perirhinal cortex is the region situated within the medial temporal lobe and is involved in visual perception, memory, and several types of learning [7778]. Ablation studies in perirhinal cortex of rats and primates show impairment in recognition memory tasks; however, such studies on the hippocampal lesions showed less effect [79808182]. Perirhinal cortex is composed of agranular cortex with five layers and dysgranular cortex with six layers, but the cell types within this region have not been well studied yet [83].
The perirhinal cortex showed mGluR-dependent LTD when DHPG (50 µM, 20 min) or DGC IV (1 µM, 20 min) was applied; however, Group III mGluR agonist L-AP4 (0.1~1 mM) showed only acute depression in the superficial layer (layer I and II/III) during drug application [84]. LFS (200 stimuli, 1Hz) with depolarization of postsynaptic neuron to –40 mV also induced LTD in the layer II/III neurons of perirhnial cortex [25]. This LTD was blocked by APV (50 µM), MCPG (500 µM), Group I mGluR antagonist AIDA (500 µM). The Group II mGluR antagonist EGLU (200 µM) only blocked the LTD when LFS was delivered at –70 mV but not –40 mV. Moreover, DHPG (50 µM) was not able to induce chemical LTD, but when DCG IV (0.5 µM) was treated together, LTD was induced. These results indicate the involvement of Group I and II mGluR in the two type of perirhinal cortex LTDs (Fig. 1D).
These mGluRs are also necessary in familiarity discrimination [85]. When MPEP (3 mg/kg) or Group II mGluR antagonist LY 341495 (3 mg/kg) was systemically administered at the acquisition period, it had no effect on familiarity discrimination. However, when the two drugs were combined, familiarity discrimination was impaired at 24 h, but not at 15 min after drug administration. The drugs had no effect when it was treated after the sample phase of before the test. Thus, these results address the importance of Group I mGluR, especially mGluR5, and Group II mGluR in the acquisition.
Visual cortex
The visual cortex is the region in the dorsal part of our brain responsible for visual information processing. It is one of the most well-investigated regions and is a popular subject for neuroscientific investigation [8687888990]. The visual cortex is composed of six layers and layer II/III and V are the major pyramidal cell layers [91]. Inhibitory interneurons are positioned in the layer II/III, IV and V, and it consists of PV, SOM, vasoactive intestinal peptide (VIP) and reelin positive intereneurons [92].
However, limited data are available on mGluR-dependent LTD in the visual cortex [93]. LTD was induced when quisqualate (10 µM), an agonist of the AMPAR, kainate receptor (KAR) and Group I mGluR, was introduced into the rat visual cortex in the presence of CNQX, AMPA and KAR antagonist, and APV [23]. Applying mGluR agonist trans-1-amino-cyclopentane-1,3-dicarboxylic acid (tACPD, 10 µM) itself had only an acute depression effect in layer II and III neurons of rat visual cortex slices; however, LTD was induced by the combined treatment with APV and tetanization [2324]. There was also a study examining the layer variations of LTD in the rat visual cortex using pairing protocols [26]. They treated LY 341495 (100 µM) in a high concentration to block all mGluRs and found that only LTD in layer VI neurons were mGluR-dependent with the pairing protocol. In mice, LFS-induced LTD was dependent on NMDAR and mGluR2 in layer II/III neurons [94]. LTD was induced by stimulation with 900 pulses of 1 Hz, but was blocked by treatment with APV (50 µM) or mGluR2 antagonist MCCG (100 µM, Fig. 1E). It was also impaired in mGluR2 KO mice. Chemical LTD was also induced with DCG IV (1 µM, 20 min).
However, the ocular dominance plasticity was normal in mGluR2 KO mice indicating the independence of mGluR2 dependent LTD in this process.
Go to :
CONCLUDING REMARKS
We have reviewed the mGluR-dependent LTDs in a variety of cortical regions of the brain. In addition, we provided an overview of the physiological conditions involved in this type of LTD and mGluR activation itself. The ACC and IC showed the relation of Group I mGluR-dependent LTD and pain related situations. Group II mGluR-dependent LTD was observed in the PFC and perirhinal cortex, which are involved in addiction and learning. LTD in the visual cortex was also dependent on Group II mGluR, but further investigations are necessary to determine the related functions.
Most of the reported and on-going studies on mGluR synaptic plasticity involve the role of the hippocampus, cerebellum, VTA, and striatum in learning and memory, motor function, and drug addiction fields. Neurodegenerative disease models are frequently used in these investigation. The cortex regions examined in this review have been investigated to a relatively lesser extent, and further investigations are necessary in relation to other specific behaviors or pathological situations. In addition, recent advances in pharmacology and technology are expected to facilitate studies on in vivo synaptic plasticity during such behaviors or specific conditions to identify solutions to more complex questions.
Go to :
 XML Download
XML Download