

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Vascular endothelial cells are critically implicated in the modulation of vascular tone through the production of a variety of vasoactive substances. This modulation cells is known to be impaired in pathophysiological states such as diabetes mellitus, hypertension, and hyperlipidemia [1]. Nitric oxide (NO) is a major endothelium-dependent relaxing factor, and its production by vascular endothelial cells plays a vital role in the regulation of vascular motor tone and stability of blood flow as well as blood pressure [23]. Endothelial nitric oxide synthase (eNOS), one of three NOS isoforms produces NO, which has a key role in regulating systemic blood pressure [4]. Previous reports have shown that a decrease in NO production can lead to hypertension [5], and eNOS mutation leads to impaired endothelium-dependent vasorelaxation and may have hypertensive effects [6]. Taken together, NO produced by eNOS is a fundamental determinant of cardiovascular homeostasis, and regulates systemic blood pressure, vascular remodeling and angiogenesis. Furthermore, it has been reported that eNOS is activated by direct phosphorylation of serine 1179 (Ser1179) by Akt, a downstream target of phosphatidylinositol 3-kinase [7]. NO is produced by vascular endothelial cells using L-arginine as a substrate in a process catalyzed by NOS [8] and induces vascular smooth muscle relaxation by activation of guanylate cyclase [89]. Inhibition of eNOS with Nω-nitro-L-arginine methyl ester (L-NAME), resulted in significantly enhanced blood pressure due to decreased NO production [10]. Recent studies suggest that production of NO is reduced and endothelium-dependent vasorelaxation is blunted in patients with essential hypertension [11].
Nafamostat mesilate (NM), a serine protease inhibitor, is used to treat patients suffering from disseminative blood vessel coagulation, hemorrhagic lesions, and hemorrhagic tendencies. NM also improves acute pancreatitis and prevents blood clots formation during extracorporeal circulation [1213]. Our previous studies have indicated that NM inhibits tumor necrosis factor alpha (TNF-α)-induced vascular endothelial cell dysfunction [14], and attenuates transient focal ischemia/reperfusion-induced brain injury [15]. Recent reports suggest that the anti-inflammatory mechanisms in the vascular wall, which include external anti-inflammatory signals and intracellular mediators, protect endothelial cells, in part due to NO production [1617]. Moreover, there is a diverse range of serine protease inhibitors, including synthetic chemical inhibitors, and natural proteinaceous inhibitors, available for research or therapeutic purposes. Szabo et al. reported that synthetic serine protease inhibitors have an effect on vascular relaxation after cardiac surgery [18]. Chao et al. reported that kallistatin, a natural serine protease inhibitor, is a potent vasodilator which operates via a vascular smooth muscle mechanism [19]. However, the study was limited to the detection of vascular contraction and did not investigate the associated regulation mechanisms and signaling pathways. NM, as another serine protease inhibitor, may possess a similar vasodilator ability.
Therefore, in this study, we addressed this hypothesis by studying the effect of NM on NO production, NO bioavailability and vascular relaxation in vivo and NO production, arginase activity in vitro; mediated through the Akt/eNOS phosphorylation dependent signaling pathway.
Go to : 
METHODS
Cell culture
Human umbilical vein endothelial cells (HUVECs) were purchased from Clonetics (San Diego, CA, USA) and cultured in endothelial growth medium (EGM-2). Sub-confluent, proliferating HUVECs at passages 2~8 were used.
Western blot analysis
Anti-phospho-eNOS antibody was purchased from Cell Signaling (Beverly, MA, USA). Anti-NOS3, anti-β-actin, anti-phospho-Akt and total Akt antibodies were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Western blot analysis was performed by boiling 30 µg of whole cell lysate or 30 µg of tissue homogenate (obtained from rat aorta) in sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS–PAGE) loading buffer, before separation by electrophoresis and transfer to a nitrocellulose membrane. After incubation in appropriate primary and peroxidase-conjugated secondary antibodies (Santa Cruz Biotechnology, Santa Cruz, CA, USA), chemiluminescent signaling was developed using Super Signal West Pico or Femto Substrate from Thermo Fisher Scientific (Pierce, Rockford, IL, USA). Blots were imaged and band densities quantified with a Gel Doc 2000 Chemi Doc system using Quantity One software from Bio-Rad (Hercules, CA, USA). Values were normalized to a β-actin loading control.
Animals
The present study utilized 6 week old male Sprague–Dawley (SD) rats (Samtako, Osan, Korea) with body weights of between 250 and 280 g. All experimental procedures adhered to the policies of Chungnam National University regarding the use and care of animals. All animals were housed in a standard environment with a 12:12 h light/dark cycle, a constant room temperature maintained at 20~25℃, and 40~60% humidity. Food and water were supplied ad libitum.
Nitrite and nitrate measurements
Two NO metabolites, nitrite (NO2–) and nitrate (NO3–), the stable breakdown products of NO, were quantified using a commercially available Nitrate/Nitrite Fluorometric Assay Kit from Cayman Chemicals (Lexington, KY, USA), as per the manufacturer's instructions. Plasma obtained from the rat blood was deproteinized using a 10 kDa cutoff filter (Microcon YM-10, Millipore, USA) and used for the quantification of NO.
DAF-FM DA staining
DAF-FM DA is a cell-permeable fluorescent probe for the detection of NO. This was chosen in preference to the widely used DAF-2 as DAF-FM and DAF-FM DA are more sensitive to NO, more photo-stable, and less pH sensitive. DAF-FM DA permeates living cells and is rapidly transformed into water-soluble DAF-FM by cytosolic esterases. Aortic sections stained with DAF-FM DA were imaged using a fluorescence microscope. All images from control and NM treated rings were captured using identical laser intensity, brightness, and contrast settings.
Arginase activity assay
Arginase activity was measured using Quanti Chrom Arginase Assay Kit (DARG-200) from Bioassay systems (Hayward, CA, USA) following the manufacturer's instructions.
Vascular reactivity
Rats were sacrificed via sodium pentobarbital overdose. A mid-sternal split was quickly performed, and the descending thoracic aorta was carefully excised and placed in ice-cold Krebs buffer (118.3 mM NaCl, 4.7 mM KCl, 2.5 mM CaCl2, 1.2 mM KH2PO4, 25 mM NaHCO3, 1.2 mM MgSO4, 11 mM glucose, and 0.0026 mM EDTA-CaNa 2). The aorta was cleaned of excess fat, cut transversely into 5~10 rings (2.0~3.0 mm), and maintained at 37℃ and pH 7.4. Endothelium-dependent vasodilation was determined by generating dose–response curves in aortic rings pre-constricted with phenylephrine.
Statistical analysis
All experiments were performed at least three times. All data are expressed as means±standard deviations. Statistical analysis was performed using Sigma Stat (Systat Software, La Jolla, CA, USA). Data in which two conditions were compared were evaluated using one-way analysis of variance followed by Tukey's post-hoc test; p-values<0.05 were considered significant.
Go to :
RESULTS
NM stimulates p-eNOS and p-Akt expression in HUVECs
We investigated the effect of NM on the expressions of p-endothelial cell nitric oxide synthase (p-eNOS) and p-Akt in endothelial cells. For the dose-dependent experiment, endothelial cells were treated with different doses of NM for 2h and harvested for western blot analysis. For the time-dependent experiment, HUVECs were treated with 300 ng/ml of NM for different time periods from 5 min to 2 h before analysis by western blot. As shown in Fig. 1A and 1D, phosphorylation of eNOS and Akt increased in both dose- and time-dependent manner, compared to the total forms of the respective proteins. These results suggest that NM stimulates p-eNOS and p-Akt expression in endothelial cells.
 | Fig. 1Nafamostat mesilate (NM) dose- and time-dependently stimulates phosphorylation of endothelial nitric oxide synthase (eNOS) and Akt in human umbilical vein endothelial cells (HUVECs).(A) Dose dependent effect of NM on eNOS and Akt phosphorylation in HUVECs. Cells were treated with various concentrations (10~1000 ng/ml) of NM for 2 h then harvested for western blot analysis of p-eNOS and p-Akt. The levels of p-eNOS and p-Akt were quantified by densitometric analyses (B and C). (D) Time dependent effect of NM on eNOS and Akt phosphorylation in HUVECs. Cells were treated with 300 ng/mL of NM for various time points starting from 5 min up to 2 h followed by harvesting the cells for western blot analysis. Total forms of the proteins were included as a loading control. The levels of p-eNOS and p-Akt were quantified by densitometric analyses (E and F). All western blots are representative of three independent experiments. The data are presented as the means±SEM of three independent experiments. *p<0.05 compared with control cells.
|
NM stimulates the production of NO in HUVECs
NO is an endothelium derived relaxing factor, which plays an important role in the control of vascular tone and function. Phosphorylation of eNOS and Akt leads to stimulation of NO production in endothelial cells. We therefore measured the level of NO produced by HUVECs treated with different doses of NM. Fig. 2A shows that NM treatment dose-dependently increased the production of NO in HUVECs.
 | Fig. 2Nafamostat mesilate (NM) stimulates the production of nitric oxide and inhibits arginase activity in human umbilical vein endothelial cells (HUVECs).(A) NO production in HUVECs treated with various concentrations (10~1000 ng/ml) of NM for 2 h. (B) Arginase activity measured in HUVECs treated with various concentrations (30~1000 ng/mL) of NM for 2 h. Bars represent means±standard error (n=3).
|
NO is produced from L-arginine by the activity of eNOS. L-arginine is a common substrate for NOS, metabolized by arginase to produce urea and ornithine. We measured the arginase activity in HUVECs treated with NM and found that there was a subsequent dose-dependent decrease in arginase activity (Fig. 2B), indicating increased eNOS activity after NM treatment.
NM stimulates p-eNOS and p-Akt expression ex vivo
To identify the physiological relevance of NM on eNOS and vascular function, we first examined whether the phosphorylation of NOS and Akt in aortic rings treated with NM was activated. Aortas from SD rats were obtained, cut into segments and incubated with various doses of NM for 2 h, then analyzed by western blot analysis. As shown in Fig. 3A, phosphorylation of both eNOS and Akt increased with increasing dose of NM, similar to the in vitro results.
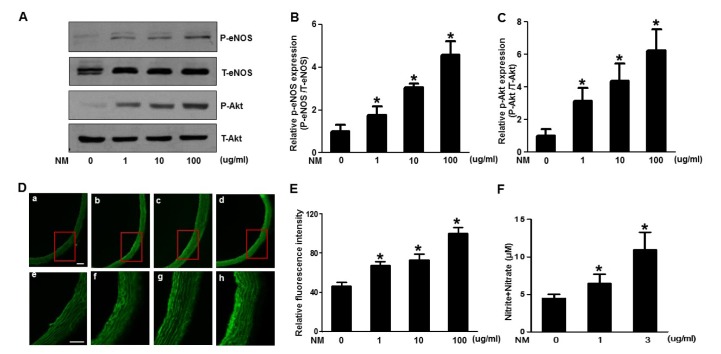 | Fig. 3Nafamostat mesilate (NM) stimulates phosphorylation of Akt/endothelial nitric oxide synthase (eNOS) and production of nitric oxide (NO) ex vivo.(A) Aorta segments were treated with various concentrations (1~100 µg/ml) of NM for 2 h then harvested for western blot analysis of p-eNOS and p-Akt. Total forms of the proteins were included as a loading control. The levels of p-eNOS and p-Akt were quantified by densitometric analyses (B and C). All western blots are representative of three independent experiments. The data are presented as the means±SEM of three independent experiments. *p<0.05 compared with the control. (D) Aorta segments were treated with various concentrations (1~100 µg/ml) of NM for 2 h, stained with DAF dye, and analyzed by microscopy. Images (a, b, c, d) were captured using 100 × magnification. Images (e, f, g, h) were captured using 200 × magnification. Scale bar=40 µm. Corresponding graphs for the relative fluorescence intensity are represented in (E). (F) Rats were treated intravenously with various concentrations (1~3 µg/ml) of NM for 2 h then measured NO production in the plasma. Bars represent means±standard error (n=3).
|
NM stimulates the production of nitric oxide ex vivo
Next, we examined the effect of NM on NO production in aortic rings. Aortas from SD rats were obtained, cut into segments and incubated with various doses of NM for 2 h followed by incubation with DAF dye for 15 min in the dark. The aortic rings were then sliced and mounted on a glass slide for analysis by microscopy. As shown in Fig. 3D, NM treatment dose dependently increased the production of NO, as evidenced by increasing DAF fluorescence intensity which is consistent with the in vitro results. We also measured NO levels in the plasma obtained from the blood of rats treated with different dose of NM. Increased dose of NM stimulated the production of NO (Fig. 3F).
NM improves impaired endothelial-dependent vascular relaxation ex vivo
Finally, we tested whether NM treatment stimulated vascular reactivity. Fig. 4A shows that NM treatment in aortic rings preconstricted with phenylephrine (10-5M) dose-dependently induced an increase in vasorelaxation. However, NM treatment did not induce as much vasorelaxation in L-NAME (which is a NOS inhibitor) pretreated aortic rings suggesting the importance of eNOS activation in the development of relaxation of vessels. At the same time, bioavailability of NO was higher in NM treated aortic rings compared to the saline vehicle control (Fig. 4B) further supporting the concept of eNOS activation. These data collectively suggest that vascular function, as well as eNOS activity, is improved with NM treatment of vascular endothelial cells.
 | Fig. 4Nafamostat mesilate (NM) improves impaired endothelial-dependent vascular relaxation ex vivo.(A) Aortic rings incubated with various concentrations (10~30 µg/ml) of NM were used to measure vascular reactivity. Increasing aortic relaxation induced by increasing dose of NM in aortic rings preconstricted with phenylephrine (10–5 M) compared with NM plus L-NAME treated aortic rings. *p<0.05compared with the control. (B) Increased bioavailable NO with NM treatment compared to saline control. Bars represent means±standard error (n=3). (C) Graphical representation of NM activity and pathway.
|
Go to :
DISCUSSION
In the present study, we investigated the effect of NM on NO production and vascular relaxation mediated by phosphorylation of Akt and eNOS and found that NM stimulated Akt/eNOS phosphorylation and NO production in a dose- and time-dependent manner (Fig. 4C). NM also caused a decrease in arginase activity. The NM-induced decreases in arginase activity could improve the available arginine substrate for NO production. We also detected that the aortic relaxation was significantly decreased when adding L-NAME to inhibit eNOS phosphorylation. These results suggest that NM promotes endothelium-dependent vasorelaxation by enhancing Akt and eNOS phosphorylation and that NM administration has the potential to stabilize blood pressure and may be effective to prevent cardiovascular diseases.
We previously showed that NM produces anti-inflammatory effects on vascular endothelial cells [14], and neuroprotective effects in the injured brain by inhibiting endoplasmic reticulum stress [15]. As previously reported, anti-inflammatory mechanisms protect endothelial cells partly through NO production [1719], which may be effective in NO-induced vasodilation. In the present study, relaxation of aortic rings was observed in the group which was administered NM (Fig. 4A). Subsequently, we investigated the signaling pathway responsible for vascular relaxation response to NM.
NO, the endothelium-derived vasoactive factor, plays an important role in modulating vasodilation. eNOS, also known as nitric oxide synthase 3, generates NO in blood vessels and is involved in the regulation of vascular function. Recent reports have suggested that in response to a variety of stimuli, efficient NO production requires eNOS phosphorylation via the Akt pathway [72021]. Therefore, we investigated whether NM activated Akt/eNOS phosphorylation. Previously, we have shown that incubation with various concentrations of NM (0.01~100 ug/ml) for 24 h did not affect the viability of HUVECs [14]. In the present study, our data showed that NM (10~1000 ng/ml) treatment induced Akt and eNOS phosphorylation in a dose- and time-dependent manner in HUVECs within 2 h (Fig. 1), suggesting that NM-induced Akt phosphorylation mediates the activation of eNOS, and may lead to increased NO production. Next, we measured the downstream target of Akt/eNOS phosphorylation that is NO production (Fig. 2). Another possibility is that NM induces vasorelaxation directly, not via activation of Akt/eNOS pathway and subsequent increased NO production. In order to clarify the mechanism, we added L-NAME to inhibit eNOS activity. The aortic relaxation was significantly decreased (Fig. 4A), demonstrating that NM-induces vasorelaxation fully, or in part, via eNOS-mediated NO production. NO is produced from L-arginine by eNOS via the L-citrulline/L-arginine recycling pathway [22]. The enzyme arginase metabolizes L-arginine to L-ornithine and urea. Increasing evidence suggests that arginase upregulation functionally inhibits available arginine substrate for NOS activity and contributes to decreased NO generation [2324], which means that decreased arginase activity increases l-arginine levels, the common substrate for NOS. Therefore, NM-induced depressed arginase activity observed in this study may promote NO production by releasing more available arginine substrate (Fig. 2B).
Previous studies have demonstrated that NM stimulates Akt/eNOS phosphorylation and NO production in a dose- and time-dependent manner in HUVECs. To examine more specifically the role of NM on endothelium-dependent vasodilation, we detected Akt/eNOS phosphorylation and NO production in rat aortic tissues. Our data showed that NM increased Akt/eNOS expression in a dose-dependent manner in rat aortic tissues (Fig. 3A), as well as NO production in plasma (Fig. 3F). We also observed a significant amount of Akt and eNOS phosphorylation in aortas stimulated with high concentration of NM, nearly 100 times the concentration required in HUVECs. We have clearly observed that NM evoked a conclusive increase in NO-related fluorescence of aorta tissues stained with the fluorescent probe DAF-FM-DA (Fig. 3D). Therefore, it is now clear that NM is able to directly activate NO production in the vessels. However, pretreatment with L-NAME significantly inhibited NM-induced vasodilatation to basal levels, suggesting that eNOS activation may be the major cause of NO production in the aorta. Overall, we can conclude that NM-induced Akt/eNOS-phosphorylation promotes NO production and NM-induced decrease in arginase activity improves the available arginine substrate for NO production and this released NO promotes endothelium-dependent vascular relaxation (Fig. 4C). These data are consistent with findings of the vasorelaxation induced by other serine proteases inhibitors. Although our findings further elucidate signaling pathways of NM-induced vasorelaxation, it is not clear how NM receptors activate Akt. Therefore, further studies are required to elucidate how NM receptors transmit signals to the Akt/eNOS pathway.
In summary, NM stimulates endothelium-dependent vascular relaxation. This vascular effect is mediated by increased production of NO, which occurs via the activation of eNOS via Akt/eNOS signaling pathway.
Go to :
 XML Download
XML Download