

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Inflammation is one of the innate immunity barriers involved in the process of removing pathogens [1]. This response includes numerous molecular and cellular reactions such as the production of cytokines, chemokines, inflammatory mediators, and complement proteins by modulating NF-κB- or AP-1-mediated transcriptional activation pathways [23]. The activation of inflammatory responses also increases the movement of blood leukocytes into inflamed areas by adhesion and chemotaxis processes [4]. These events require functional activation of adhesion molecules triggered by chemokines and ligation with counter-ligands when cells homotypically and heterotypically interact with leukocytes and endothelial cells [5].
Monocytes are important cells in inflammatory responses because blood monocytes are needed to supply fresh macrophages after full differentiation [6]. Macrophages are phagocytes which play a critical role in removing microorganisms [7]. Therefore, stimulation of monocyte migration could help to boost inflammatory responses by increasing numbers of tissue-resident macrophages. The migration activity of monocytes is managed by adhesion molecule activity. So far, variety of adhesion molecules are known to participate in the activation of monocytes. Previously, we and others have identified that β1-integrin (CD29), CD98, CD147, and CD43 are major functional adhesion molecules expressing in monocytes. The activation of these molecules by ligation with homotypic or heterotypic ligands is reported to induce intracellular signaling pathways leading to functional activation of monocytes playing a critical roles in inflammation and virus-derived fusion events [8]. Activation signals of monocytes induced by these adhesion molecules include small GTPase Rho, tyrosine kinases (Syk and Lyn), and the phosphatidylinositol-3-kinase during CD29 activation [910], ERK, Syk, and protein kinase Cθ in CD43 activation [1112], conventional PKC isoforms (α, β, γand δ), ERK, and p38 in case of CD98 stimulation [111314], and VEGFR-2 tyrosine kinase receptor, PI3K, AKT, and ERK1/2 under CD147 activation conditions [151617].
Of them, CD29 is regarded as a significant adhesion molecule that is critically important in allowing strong interactions between leukocytes and endothelial cells in the process known as extravasation [18]. Previously, we and other groups have found that CD29 is functionally associated with CD98 [1920]. Moreover, it was found that CD98 is regulated by CD147 [2122]. Although several reports have suggested cross-regulation between CD98 and CD147 [23], detailed mechanisms of their molecular interactions have not been fully elucidated yet. These findings raise the hypothesis that these molecules might be important for the functional activation of CD29. In this study, we aimed to further clarify the regulatory mechanisms between these three adhesion molecules at the molecular level in order to understand how CD29 is regulated by other adhesion molecules.
Go to : 
METHODS
Materials
Enzyme inhibitors [U0126, an MEK1 inhibitor, rottlerin, a protein kinase Cd inhibitor, and cytochalasin B (Cyto B), an actin cytoskeleton disruptor] were purchased from Calbiochem (La Jolla, CA, USA). U937 cells, a human pleura/pleural effusion promonocyte-like cell line (no. CRL-1593.2), were obtained from the American Type Culture Collection (ATCC, Rockville, MD, USA). All other chemicals were of reagent grade. The following antibodies were used in this study for cell-cell adhesion assays: CD29 (MEM 101A, IgG1, ascites, kindly provided by V. Horejsi); CD43 (161-46, ascites, IgG1, kindly provided by R. Villela); CD98 (ANH-18, purified IgG1, kindly provided by K. Skubiz); and CD147 (MEM M6/1, IgG1, ascites, V. Horejsi). Rhodamine phalloidin was purchased from Molecular Probes (Carlsbad, CA, USA). Antibodies to CD98 (mouse, 4F2) and CD147 (rabbit, EMMPRIN) for immunoprecipitation and immunoblotting were from Santa Cruz Biotechnology (Santa Cruz, CA, USA) and Sino Biological Inc. (Beijing, China), respectively. The following antibodies were used for flow cytometic analysis: CD18 (BU86, IgG1, ascites); CD29 (MAR4, IgG1, ascites); CD43 (161-46, IgG1, ascites); CD44 (E1/2, purified IgG1); CD98 (BK19.9, IgG1, purified antibody); and CD147 (MEM M6/1, IgG1, ascites), as reported previously [21].
Cell culture
U937 cells were cultured in RPMI1640 supplemented with antibiotics (100 U/ml of penicillin and 100 µg/ml of streptomycin) and 10% FCS, and were maintained at 37℃ and 5% CO2 under humidified air conditions.
Flow cytometric analysis
Expression of U937 surface adhesion molecules was determined by flow cytometric analysis as reported previously [2124]. Cells (105) were washed with staining buffer (PBS containing 2% FBS and 1% sodium azide) and incubated in 50 ml staining buffer containing 10% rabbit serum for 10 minutes on ice and then with the primary antibodies to CD18, CD29, CD43, CD44, CD82, CD98, and CD147 for a further 45 minutes. After washing 3 times with staining buffer, cells were treated with 1/20 dilution of fluorescein isothiocyanate (FITC)–conjugated rabbit anti–mouse IgG secondary antibody (Dako, Dakopatts, High Wycombe, United Kingdom). Cells were then washed 3 times with staining buffer and analyzed on a FACScan (Becton Dickinson, San Jose, CA).
Quantitative cell-cell and cell-fibronectin adhesion assays
A quantitative homotypic aggregation assay was carried out as reported previously [21]. Briefly, U937 cells were preincubated with enzyme inhibitors or aggregation-blocking antibodies at 37℃ for 3 h or 6 h and incubated further with aggregative antibodies (1 µg/ml) for 3 to 6 h in round-bottom microwell plates. The numbers of non-aggregated and total cells were counted in a hemocytometer and the percentage of cells in aggregates determined as 100 × (total cells–non-aggregated cells)/total cells, and referred to a control (as a percentage) calculated from only the aggregation-activating antibody treatment group. Inhibitory effects of aggregation-blocking antibodies or enzyme inhibitors were calculated as % of control by setting the control group as 100%. For the cell–fibronectin (FN) adhesion assay, drug (U0126 or Cyto B) or blocking antibody (P5D2)-treated U937 cells (5×105 cells/well) were seeded on a fibronectin (50 µg/ml)-coated plate and incubated for 3 h, as reported previously. After removing unbound cells with PBS, the attached cells were treated with 0.1% crystal violet for 15 min. The OD value at 540 nm was measured by a Spectramax 250 microplate reader.
Phase-contrast microscopy
Images of cells in culture were observed using an inverted phase-contrast microscope attached to a video camera, and captured using NIH image software as reported previously [25].
Confocal microscopy
Confocal analysis of actin and adhesion molecules (CD29, CD98, and CD147) on the membrane of U937 cells (1×105 cells/ml) treated with aggregation-activating antibodies was carried out on glass-bottom culture dishes (MatTek Corp., Ashland, MA, USA) using FITC- or PE-labeled antibodies to CD29, CD147, or CD98, as well as the actin-staining chemical rhodamine phalloidin. Intensity changes in CD29, CD147, CD98, or actin cytoskeleton were imaged by LSCM on a Zeiss LSM 510 META confocal microscope equipped with a Zeiss 37℃ incubation system. Images were analyzed using the Zeiss LSM Image Examiner.
Immunoblotting and immunoprecipitation
Cells (5×106 cells/ml) were lysed in lysis buffer (in mM: 20 TRIS-HCl, pH 7.4, 2 EDTA, 2 EGTA, 50 β-glycerophosphate, 1 sodium orthovanadate, 1 dithiothreitol, 1% Triton X-100, 10% glycerol, 10 µg/ml leupeptin, 10 µg/ml aprotinin, 10 µg/ml pepstatin, 1 benzimide, and 2 hydrogen peroxide) for 30 min under rotation in a cold room. The lysates were clarified by centrifugation at 16,000 g for 10 min at 4℃. Soluble cell lysates were immunoblotted and CD98 and CD147 were visualized as reported previously [26]. For immunoprecipitation, cell lysates containing equal amounts of protein (500 µg) from ANH-18-treated U937 cells (1×107 cells/ml) were pre-cleared with 10 µl of protein A-coupled Sepharose beads (50% v/v) (Amersham, UK) for 1 h at 4℃. Pre-cleared samples were then incubated with 5 µl of antibody to CD147 or CD98 overnight at 4℃. Immune complexes were mixed with 10 ml of protein A-coupled Sepharose beads (50% v/v) and incubated for 3 h at 4℃.
Statistical analysis
All values are presented as means±standard deviations (SDs) of three (Fig. 1, 4C, and 5B) or six (Fig. 2A, 3A, and 4A) experiments. For statistical comparisons, results were analyzed using ANOVA/Scheffe's post hoc test and the Kruskal–Wallis/Mann–Whitney tests. A p-value<0.05 was considered statistically significant. All statistical tests were performed using SPSS software (SPSS Inc., Chicago, IL, USA). Similar results were obtained in a second experiment.
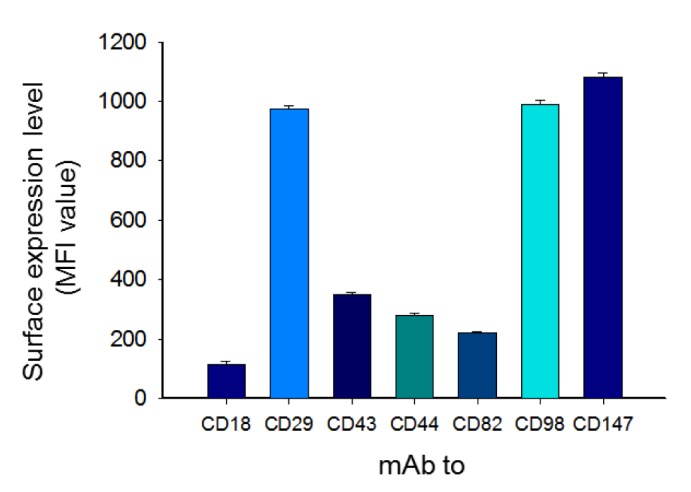 | Fig. 1Flow cytometric analysis of surface adhesion molecule expression in U937 cells.Surface adhesion molecules [CD29 (β1-integrins), CD43, CD98, and CD147] in U937 cells (1×105) were analyzed by flow cytometry. All staining antibodies were used at 1~5 µg/ml. Mean fluorescence intensity (MFI, percent of control) was calculated using WIN-MDI software from a minimum of 7,500 cells. The results (mean±SD, n=4) show one representative experiment out of three.
|
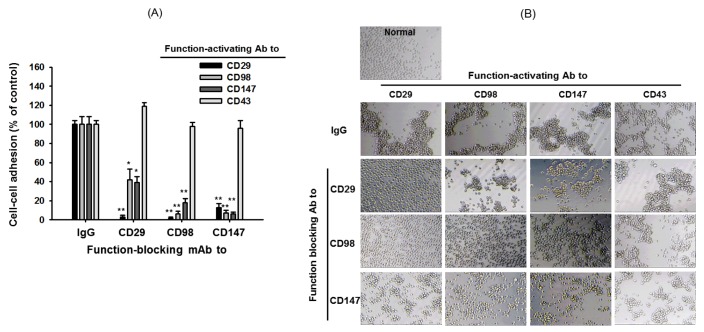
 | Fig. 4Effect of pharmacological inhibitors of ERK, PKCδ, and actin polymerization on cell-cell aggregation or cell-fibronectin adhesion induced by ligation of surface adhesion molecules with aggregation-activating antibodies or immobilized fibronectin.(A left panel) U937 cells were incubated with pro-aggregative (activating) antibodies (1 µg/ml each as IgG1) to CD98 (AHN-18, 1 µg/ml), CD29 (MEM101A, 1 µg/ml), CD147 (M6-1D4, 10 µg/ml), and CD43 (161-46, 1 µg/ml) in the presence of chemical inhibitors to ERK (U0126, 25 µM), PKCδ (rottlerin, 10 µM), and actin polymerization (Cyto B: cytochalasin B, 10 µM) for 3 h. Aggregation of cells in the absence of stimuli (normal conditions) was less than 4%. Percentage of aggregation was quantitatively determined by cell-cell adhesion assays. (A right panel) Images of the aggregated cells in culture were obtained using an inverted phase-contrast microscope attached to a video camera, and captured using NIH image software. (B) U937 cells pretreated with 10 µg/ml of function blocking antibody to CD29 (P5D2) or inhibitors [U0126 (20 µM) and cytochalasin B (10 µM)] were seeded on fibronectin (50 µg/ml)-immobilized plates and further incubated for 3 h. Attached cells were determined by crystal violet assay, as described in Materials and Methods. (C) Cytotoxic activity of U0126, rottlerin, and CytoB was confirmed by a conventional MTT assay. Results (aggregation relative to control culture in the presence of stimuli or % of cytotoxicity) are expressed as mean±SEM from three independent experiments performed in triplicate. **p<0.01 compared to the control group.
|
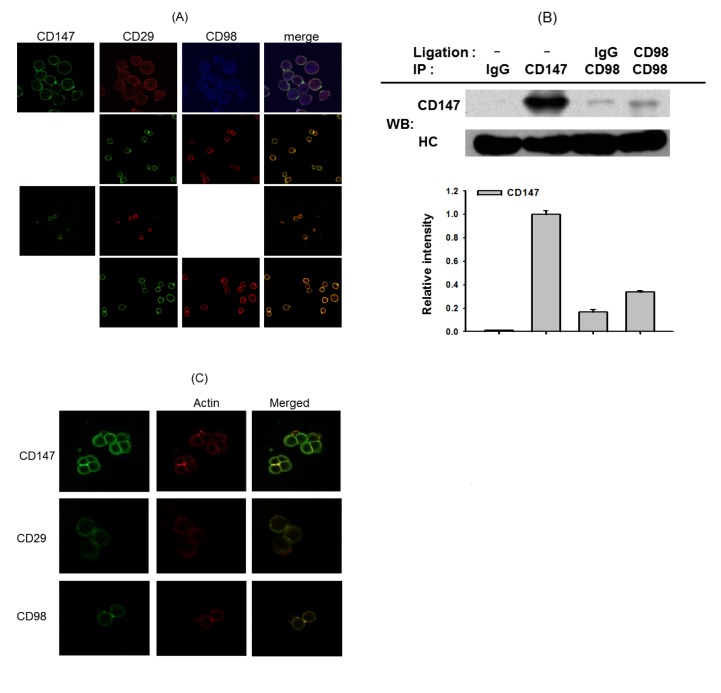 | Fig. 5Molecular association between CD29, CD98, CD187, and the actin cytoskeleton in U937 cell-cell adhesion.(A and C) U937 cells were incubated with pro-aggregative (activating) antibodies (1 µg/ml each as IgG1) to CD98 (AHN-18, 1 µg/ml), CD29 (MEM101A, 1 µg/ml), CD147 (M6-1D4, 10 µg/ml), and CD43 (161-46, 1 µg/ml) for 3 h. The localization patterns between CD29, CD98, CD147, and the actin cytoskeleton were evaluated by confocal microscopy. Results show one experiment out of three. (B) U937 cells (1×107 cells/ml) were treated with antibodies to CD147 (M6-1D4) and CD98 (AHN-18) for 6 h. After immunoprecipitation with antibodies to CD147 or CD98, the level of co-immunoprecipitated CD147 was determined by immunoblotting analysis with anti-CD147 antibodies. Results represent one experiment out of three. Relative intensities were calculated with the DNR Bioimaging system (Gelquant software Ver. 2.7).
|
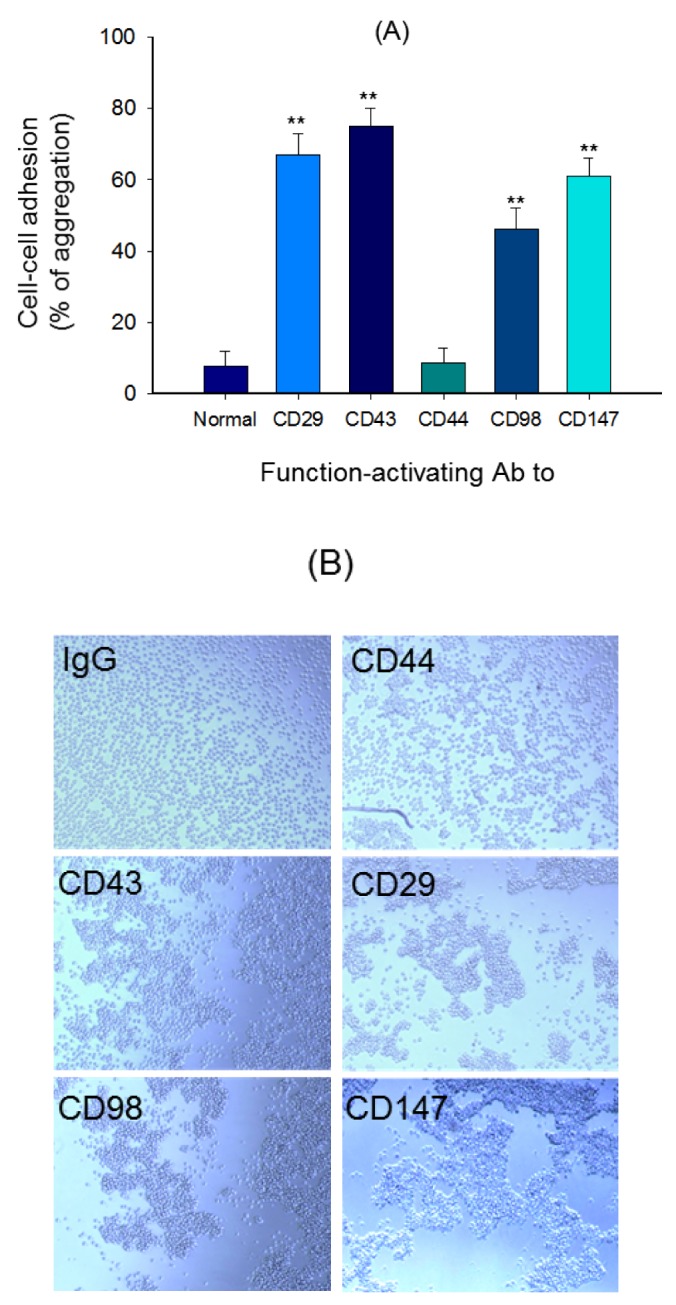 | Fig. 2Cell-cell adhesion induced by ligation of surface adhesion molecules with aggregation-activating antibodies.(A) U937 cells were incubated with pro-aggregative (aggregation-activating) antibodies (1 µg/ml each as IgG1) to CD98 (AHN-18, 1 µg/ml), CD29 (MEM101A, 1 µg/ml), CD147 (M6-1D4, 10 µg/ml), and CD43 (161-46, 1 µg/ml) for 6 h. Aggregation of cells in the absence of stimuli (normal conditions) was less than 4%. Percentage of aggregation was quantitatively determined by cell-cell adhesion assays. (B) Images of the aggregated cells in culture were obtained using an inverted phase-contrast microscope attached to a video camera, and captured using NIH image software. Results (aggregation relative to control culture in the presence of stimuli) are expressed as mean±SEM from three independent experiments performed in triplicate. **p<0.01 compared to the control group.
|
 | Fig. 3Effect of blocking antibodies on cell-cell adhesion induced by ligation of surface adhesion molecules with aggregation-activating antibodies.(A) U937 cells were incubated with pro-aggregative (aggregation-activating) antibodies (1 µg/ml each as IgG1) to CD98 (AHN-18, 1 µg/ml), CD29 (MEM101A, 1 µg/ml), CD147 (M6-1D4, 10 µg/ml), and CD43 (161-46, 1 µg/ml) in the presence of aggregation-blocking antibodies to CD29 (P5D2, 1 µg/ml), CD98 (MEM 108, 1 µg/ml), and CD147 (MEM M6/1, 1 µg/ml) for 6 h. Aggregation of cells in the absence of stimuli (normal conditions) was less than 4%. Percentage of aggregation was quantitatively determined by cell-cell adhesion assays. (B) Images of the aggregated cells in culture were obtained using an inverted phase-contrast microscope attached to a video camera, and captured using NIH image software. Results (aggregation relative to control culture in the presence of stimuli) are expressed as mean±SEM from three independent experiments performed in triplicate. *p<0.05 and **p<0.01 compared to the control group.
|
Go to :
RESULTS
Expression levels of CD29, CD43, CD98, and CD147 and effect of their ligation with aggregation-activating antibodies
To check expression levels of surface adhesion molecules in U937 cells, flow cytometric analysis was employed. As Fig. 1 shows, the MFI values of CD147, CD98, CD29, and CD43 were 1140, 1013, 992, and 386, respectively. The lowest expression level was seen in CD18 (Fig. 1). Similar to expression levels, aggregation-inducing activity of U937 cells was also exhibited in the ligation of CD43, CD29, CD147, and CD98 up to 76, 68, 61, and 48% compared to 9% of normal cells (Fig. 2A and 2B).
Effect of blocking antibodies and pharmacological inhibitors on cell-cell adhesion induced by the activation of CD29, CD43, CD98, and CD147
Whether the function-blocking antibodies to CD29, CD98, and CD147 are able to interrupt the cell-cell adhesion induced by the activation of CD29, CD43, CD98, and CD147 was examined. Interestingly, P5D2, a blocking antibody to CD29, blocked the aggregation events induced by CD29, CD98, and CD147 up to 92, 57, and 62%, respectively, while MEM 108 (a CD98 blocking antibody) also suppressed the aggregation levels up to 94, 89, and 78%, respectively (Fig. 3A and 3B). Similarly, the adhesion events induced by CD29, CD98, and CD147 were inhibited by CD147 blocking antibody MEM M6/1 up to 82, 86, and 90%, respectively (Fig. 3A and 3B). However, there was no significant inhibition of blocking antibodies in CD43-triggered cell aggregation (Fig. 3A and 3B).
In addition, sensitivity of cell-cell adhesion events to several enzyme inhibitors was also tested under the aggregation-inducing conditions. Thus, U0126, a MEK1 inhibitor, displayed strong suppressive activity toward U937 cell-cell adhesion events boosted by CD29, CD98, and CD147 up to 92, 84, and 82%, respectively (Fig. 4A). Rottlerin, a PKCδ inhibitor, also diminished the aggregation events by 90, 86, and 72%, whereas Cyto B, an actin cytoskeleton disruptor with blocking activity on actin polymerization, almost completely inhibited the cell-cell adhesion processes managed by CD29, CD98, and CD147 up to 98%. Like blocking antibody effects, these three inhibitors failed to block CD43-induced U937 cell aggregation (Fig. 4A). Interestingly, the inhibitory levels of U0126 and Cyto B in FN-mediated U937 cell adhesion events were dramatically changed exhibiting not only marginal suppression (30%) in U0126 treatment, but also strong stimulation (170%) in Cyto B exposure (Fig. 4B). Meanwhile, no dramatic inhibition (8, 3, and 14%) of U937 cell viability by U0126, rottlerin, and Cyto B was also observed (Fig. 4C).
Molecular association between adhesion molecules (CD29, CD98, and CD187) and actin cytoskeleton
To check whether these molecules are also cellularly and biochemically associated, we employed confocal and immunoprecipitation analyses. According to confocal analysis, there was clear associated between CD147, CD29, and CD98, which are appeared on the membrane of U937 cells (Fig. 5A). In addition, by immunoprecipitation and immunoblotting analyses, binding property of CD147 to CD98 was clearly observed (Fig. 5B). Finally, tight association between actin cytoskeleton and CD147, CD98 or CD29 was also found by confocal microscopy (Fig. 5C).
Go to :
DISCUSSION
The adhesion of monocytes is one of the important steps in supplying additional macrophages during the inflammatory response [29]. Among adhesion molecules, β1-integrins are known to be functionally important [30]. In this study, we focused on understanding the regulatory mechanism of CD29 in terms of its associated co-adhesion molecules.
Fig. 1 shows the importance of CD29 and its regulatory proteins in U937 promonocytic cells. The surface levels of CD29, CD98, and CD147 were found to be high, while others (CD43, CD44, and CD82) displayed low levels on the surface. These data suggest that these three molecules might act as major adhesion molecules in monocytes. In fact, ligation of these three molecules with aggregation-activating antibodies strongly induced homotypic U937 cell-cell adhesion events, as assessed by quantitative and qualitative approaches (Fig. 2A and 2B). As reported previously, CD43 activation also led to homotypic cell adhesion patterns in U937 cells.
Aggregation-blocking antibodies to CD29, CD98, and CD147 (but not CD43) almost completely blocked each cell-cell aggregation pattern generated by CD29, CD98, or CD147 activation conditions (Fig. 3A and 3B), indicating that these molecules are closely and functionally associated. The sensitivity of cell-cell adhesion to treatment with inhibitors of signaling enzymes showed very similar patterns for CD29-, CD98-, and CD147-induced conditions, while CD43-induced aggregation was not blocked by these inhibitors (Fig. 4A and 4B). There was no cytotoxicity under treatment with these compounds (Fig. 4C), implying that the inhibitory activity of these compounds was not due to cell death.
Biochemical association between CD29, CD98, and CD147 was examined using confocal and immunoprecipitation analyses. As Fig. 5A shows, these three molecules appeared to be focally associated on the membrane of U937 cells. The molecular interaction between CD147 and CD98 was also confirmed by immunoprecipitation analysis (Fig. 5B). This interaction increased under CD98 ligation conditions, indicating that the active form of CD98 could better associate with CD147. Moreover, molecular association between CD29 and CD98 as well as between CD29 and CD147 has been also previously demonstrated by employing immunoprecipitation analysis [3132]. Since actin is known as an adaptor molecule that allows intracellular signaling molecule (e.g. Src) association [333435], we examined whether actin is important in this complex formation. As Fig. 5C shows, actin stained with rhodamine phalloidin was detected in the focally concentrated area with CD147, CD98, or CD29, implying that these molecules might be associated with the actin cytoskeleton. Indeed, it has been reported that cytochalasin B is capable of completely suppressing the clustering formation induced by CD29 and CD98 [36]. So far, it is unclear whether actin simultaneously binds to these adhesion molecules to manage the formation of molecular complex. Therefore, relevant works regarding the functional role of actin in CD29/CD98/CD147-mediated cell-cell adhesion events will be further continued.
In terms of pathophysiology, modulation of CD29 function is regarded as a therapeutic target to treat various chronic inflammatory and autoimmune diseases [303738]. CD29 was found to be highly expressed in monocytes to interact with vascular endothelial cells and lung fibroblasts and in autoreactive T lymphocytes analyzed from autoimmune patients [39404142]. In addition, engagement of CD29 on monocytes and SLE T cells was reported to trigger focal adhesion kinase- or spleen tyrosine kinase-induced intracellular signaling cascade linked to the proliferation of T cells and secretion of chemokines from monocytes [394344], implying that CD29 may play a critical role in increasing adhesive activity of monocytes and autoreactive T cell activation. The inflammatory processes mediated by neutrophils and macrophages were also found to be critically regulated by CD29 [45]. These findings have raised the possibility that functional regulation of CD29 by small molecules or therapeutic antibodies could become a useful therapeutic strategy in treating autoimmune and chronic inflammatory diseases. We have reported that lancemaside A, 20S-dihydroprotopanaxadiol, cinnamaldehyde, ceramide, chloroquine, and cynaropicrin are regarded as good modulators of CD29-mediated cell-cell adhesion [254647]. Others have found that ibrutinib (an orally active covalent BTK inhibitor) [48], shikonin [49], and biodentine [50] can inhibit the functional activation of integrins. Although there have been numerous attempts to develop CD29-function inhibitors with small molecules, none of these drugs has been released for clinical trials. There is also no therapeutic CD29 antibody (e.g. OS2966) yet prescribed for the clinic [51]. Indirect approaches with other CD29-associated molecules, such as CD98 and CD147, could lead to acceptable methods for regulating CD29 function. Treatment with aggregation-blocking antibodies against CD98 and CD147 strongly suppressed CD29-mediated cell adhesion events (Fig. 3). Further development of therapeutic approaches against autoimmune and chronic inflammatory diseases using CD98 and CD147 will provide additional opportunities for CD29-targeted drug development.
In summary, our study demonstrated that CD29 can be regulated by its associated proteins, CD98 and CD147. Monocytic U937 cells showed similar expression patterns for CD29, CD147, and CD98, and the adhesion events induced by these three molecules exhibited similar inhibitory sensitivity to enzyme inhibitors and blocking antibodies. These molecules were found to be focally associated with the actin cytoskeleton and also co-immunoprecipitated, as shown by confocal and immunoprecipitation analyses, respectively. These results suggest that co-regulation of β1-integrins and the adhesion molecules CD98 and CD147 may significantly contribute to the adhesion of monocytic U937 cells.
Go to :
 XML Download
XML Download