

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Accumulating evidence over the last few decades suggests that in addition to classical synaptic transmission, information is transmitted between cells via the diffusion of neurotransmitters into the extracellular space and the activation of extrasynaptic receptors in the brain [12]. The activation of extrasynaptic ionotropic receptors generates slow, persistent tonic currents that offer unique mechanisms of neuronal control [34], while synaptic receptors mediate rapid, phasic excitatory (E) or inhibitory (I) postsynaptic currents (EPSCs or IPSCs, respectively). In this sense, activation of extrasynaptic N-methyl-D-aspartate receptors (eNMDARs) by glutamate in the extracellular space can evoke a persistent tonic NMDA current (tonic INMDA) [5678], while synaptic NMDA receptors are responsible for classical EPSCs. The idea that a glutamate signaling mechanism can be compartmentalized via synaptic and extrasynaptic NMDARs has been also supported bysyudies showing that synaptic and extrasynaptic NMDARs are linked to distinct, and even opposing, downstream biological actions [91011]. However, despite growing evidence that supports the presence of tonic INMDA generated by eNMDARs, the functional significance of tonic INMDA in various brain regions remains poorly understood.
In the brain, α-amino-3-hydroxy-5-methylisoxazole-4-proprionate receptors (AMPARs) are responsible for the bulk of glutamatergic EPSCs and their dynamic regulation ensures dynamic fitting of the receptor function that underlies much of the plasticity of excitatory transmission. For example, calcium influx through open ATP-gated channels leads to reduced surface AMPARs in dendrites and at synapses in the hippocampus [12], while it facilitates AMPAR function in the hypothalamic paraventricular nucleus (PVN) neurons [1314]. Given that activation of either synaptic or extrasynaptic NMDARs causes changes in intracellular calcium concentration [9], it is plausible that both synaptic and extrasynaptic receptors are involved in the regulation of AMPAR function. Despite the wealth of information regarding the role of synaptic NMDARs, little is known about the role of eNMDARs in regulating AMPAR function in the brain.
Magnocellular neurosecretory cells (MNCs), composed of vasopressin and oxytocin neurons in the hypothalamic PVN and supraoptic nucleus (SON), play a major role in fluid-balance homeostasis and reproductive function [15]. As in other brain regions, glutamate is a critical excitatory neurotransmitter in SON MNCs [16]. Glutamate-generating tonic INMDA under glial control efficiently influences neuronal activity in the magnocellular neurosecretory system. Enhanced activation of eNMDARs, with blunted glial GLT-1 clearance, contributes to increased MNC activity and hormone release during dehydration and heart failure conditions [46]. However, the functional role(s) of eNMDARs that generate tonic INMDA must be further elucidated in SON MNCs. In the present study, we demonstrated that activation of eNMDARs that generate tonic INMDA facilitates AMPAR function, resulting in enhanced AMPA-induced steady-state current in SON MNCs.
Go to : 
METHODS
Experimental animals
All animal experiments adhered to the Chungnam National University policies regarding the care and use of animals. Male Sprague–Dawley rats (60~80 g) were housed under a 12/12-h light/dark schedule. Rats were randomly divided into two groups: euhydrated (EU) and chronic dehydarated (DE) animals. The EU group was allowed free access to normal tap water, whereas DE rats exposed by 2% saline for 7 days. All rats had access to food water ad libitum except for the DE periods throughout the experiments. Plasma osmolality was measured by freezing-point depression (Fiske Associates, Norwood, MA, USA) prior to sacrifice.
Electrophysiological recordings and data analysis
Hypothalamic slices (300 µm) were obtained as previously described [17]. Brian slices containing the SON were cut using a vibroslicer (Leica VT 100s, Leica, Bensheim, Germany) and placed in a holding chamber containing standard artificial cerebrospinal fluid (aCSF) until use. Standard aCSF consists of 126 mM NaCl, 26 mM NaHCO3, 5 mM KCl, 2.4 mM NaH2PO4, 2.4 mM CaCl2, 1.2 mM MgCl2 and 10 mM glucose, pH 7.3~7.4. The medium was saturated with 95% O2 and 5% CO2.
Patch-clamp recordings were obtained using an Axopatch200-B amplifier (Axon Instruments, Foster City, CA, USA). For voltage-clamp recordings, a low Mg2+ aCSF (20 µM MgCl2) was used to facilitate NMDAR-mediated currents at a holding potential (Vholding) of –70 mV. Periods of 180 sec of synaptic activity were analyzed using the Minianalysis 6.0.3 program (Synaptosoft Inc., Decatur, GA). In some cases, recordings were also obtained at a Vholding of –40 mV or –20 mV in normal standard aCSF. Currents were recorded in the presence of picrotoxin (100 µM) to inhibit ionotropic GABA receptors, if not mentioned. Current output was filtered at 2 kHz and digitized at 10 kHz (Digidata 1322A, Axon Instruments) in conjunction with pClamp 9.2 software. Patch pipettes (3~5 MΩ) were filled with a solution containing 140 mM K-gluconate, 10 mM KCl, 10 mM HEPES, 0.5 mM CaCl2, 5 mM EGTA and 5 mM Mg2+ATP, pH 7.3.
Persistent activation of AMPARs and NMDARs was defined as the difference of holding current (Iholding) before and after application of the receptor agonists: the glutamate AMPA/kainate receptor antagonists, NBQX (2,3-dihydroxy-6-nitro-7-sulfamoyl-benzo[f]quinoxaline-2,3-dione) or DNQX (5,7-dintroquinoxaline-2,3-dione, 10 µM), and the NMDA receptor antagonist AP5 (D,L-2-amino-5-phosphonopentanoic acid, 100 µM), respectively [171819]. The Iholding was measured in 50 ms epochs of traces lacking PSCs using Mini Analysis (Synaptosoft, Decatur, GA, USA).
Series resistance was monitored throughout the experiment. Neurons with series resistance changes >15% during experiments were not included in the analysis.
Statistics
Numerical data are presented as the mean±standard error of the mean (SEM). Statistical significance of the data was determined using independent or paired Student's t-test as needed.
Go to :
RESULTS
Electrophysiological recordings were obtained from 112 SON MNCs under bright-light microscopy [20]. In a low-Mg2+ bath solution, AP5 induces an outward shift in the holding current (Iholding), named tonic INMDA, mediated by eNMDAR in SON MNCs [67]. The mean amplitude of tonic INMDA in SON MNCs was 12.1±2.5 pA (n=7) in our recording conditions (20 µM of MgCl2 and Vholding of –70 mV). In contrast, AP5 caused no significant changes in Iholding in normal aCSF containing 1.2 mM MgCl2.
Tonic activation of AMPARs by glutamate in SON MNCs
To determine whether INMDA generated by eNMDARs alters AMPARs function to change the response to its agonists, glutamate was bath applied with a known concentration of the agonist (10 µM) in low Mg2+ aCSF. Glutamate caused a steady-state inward shift in Iholding (IGLU) at Vholding of –70 mV, which was blocked by the sequential application of NBQX and NBQX+AP5 in SON MNCs (Fig. 1A and 1B). Interestingly, glutamate failed to change Iholding in the presence of AP5 in SON MNCs (Fig. 1A and 1B), suggesting that glutamate induced persistent tonic activation of AMPARs in an NMDAR-activation-dependent manner in SON MNCs. In agreement with this, glutamate induced NBQX-sensitive currents in normal aCSF when Vholding was depolarized to –40 mV or –20 mV, activating NMDARs. While glutamate caused minimal changes in Iholding (p>0.2, n=5) at Vholding of –70 mV, it induced NBQX-sensitive IGLU at THE depolarized Vholding (–40 mV, 16.6±2.9 pA, n=7; –20 mV, 14.6±4.9 pA, n=7) (Fig. 1C and 1D). Furthermore, IGLU was completely inhibited by pretreatment with AP5 at all tested potentials in SON MNCs (data not shown). In a subset of experiments, we further confirmed that IGLU was blocked by the additional application of AP5 (Fig. 1C) inset.
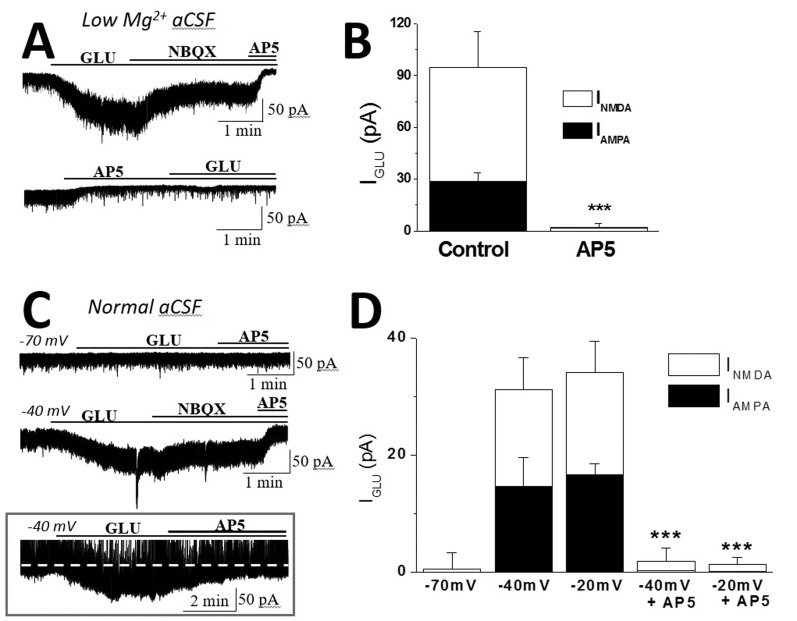 | Fig. 1Glutamate evoked NBQX-sensitive tonic currents in an NMDAR activation-dependent manner in SON MNCs.(A) Representative current traces showing that IGLU was blocked by the sequential application of NBQX (IAMPA) and NBQX+AP5 (INMDA) in low-Mg2+ aCSF. (B) The mean IAMPA and INMDA in the absence (n=7) and presence (n=3) of AP5 are summarized as in A. (C) Representative current traces showing that glutamate evoked IAMPA in an INMDA-dependent manner at depolarized holding potentials in normal aCSF. Note that IGlu was efficiently blocked by AP5 alone, which was not affected by picrotoxin (inset, Vholding of –40 mV in normal aCSF). (D) The mean IAMPA and INMDA at different holding potentials (n=3~7) are summarized as in C. ***p<0.001 compared to each control.
|
NMDARs blockade reduced AMPA-induced current in SON MNCs
To determine whether NMDAR activation caused an alteration in the steady state sensitivity of the AMPA receptor-activated channels to its agonists, AMPA was bath-applied in low-Mg2+ aCSF and normal aCSF. Application of AMPA (1 µM) induced a reproducible steady-state inward current (IAMPA) in the SON MNCs, which was blocked by an AMPAR antagonist, DNQX or NBQX (Fig. 2). We recorded and compared IAMPA amplitudes in the absence and presence of the NMDAR antagonist.
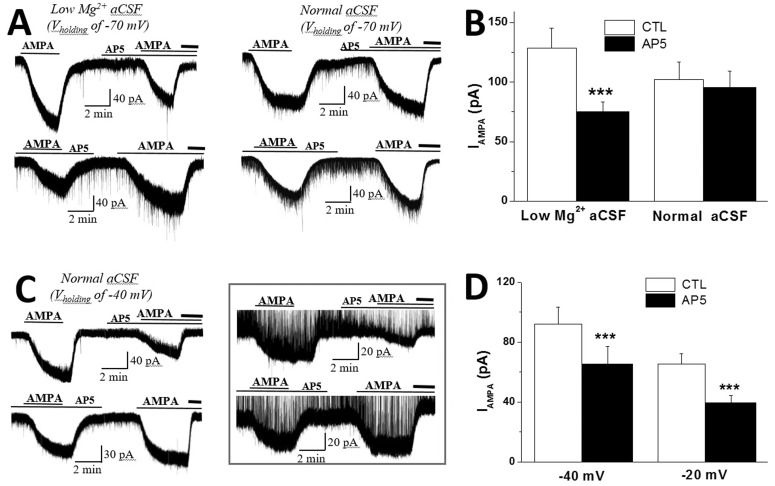 | Fig. 2NMDAR activation facilitated AMPA-induced steady-state inward currents (IAMPA) in SON MNCs.(A) Representative current traces showing IAMPA in low and normal Mg2+ aCSF (Vholding –70 mV). Note that pretreatment with AP5 consistently reduced IAMPA in low-Mg2+ aCSF, while repeated applications of AMPA increased IAMPA in normal aCSF. Bold lines at each trace represent the application of an AMPAR antagonist, DNQX or NBQX. (B) The mean IAMPA amplitudes in low-Mg2+ (n=12) and normal (n=10) aCSF are summarized as in A. Effects of AP5 on the first and second IAMPA were tested and combined from the same number of neurons in each group. (C) Representative current traces showing that the pretreatment of AP5 consistently reduced IAMPA recorded at Vholding of –40 mV in normal aCSF. IAMPA inhibition by AP5 was not affected by picrotoxin in normal aCSF (Vholding –20 mV, inset). Bold lines at each trace represent the application of AMPAR antagonist as in A. (D) The mean IAMPA amplitudes at Vholding of –40 mV (n=12) and –20 mV (n=10) are summarized. Effects of AP5 on the first and second IAMPA were pooled as in B. ***p<0.001 compared to respective controls.
|
Although there is a tendency for IAMPA amplitude to increase with repeated AMPA applications, pretreatment with AP5 consistently attenuated IAMPA amplitude in the first and second AMPA applications (Fig. 2A). The inhibitory effects of AP5 on the first and second IAMPA amplitudes were tested in the same number of neurons and combined to compare with the control values (Fig. 2B). In the presence of AP5, IAMPA amplitude was decreased to 60.3±4.06% of that in the absence of the antagonist in low-Mg2+ aCSF (control: 134.8±17.0 pA; AP5: 77.8±8.5 pA; n=12, p<0.001) (Fig. 2A and 2B), while pretreatment with AP5 failed to affect IAMPA amplitudes in normal aCSF (control: 91.7±10.9 pA; AP5: 86.9±11.5 pA; n=10, p>0.15). IAMPA facilitation by INMDA was also evident in normal aCSF at depolarized Vholding activating NMDARs (Fig. 2C and 2D). In agreement with IAMPA facilitation by INMDA in low-Mg2+ aCSF, AP5 reduced IAMPA amplitudes from 92.0±11.2 pA to 65.3±11.7 pA (n= 12, p<0.001) at Vholding of –40 mV in normal aCSF. Pretreatment with AP5 also significantly reduced IAMPA at Vholding of –20 mV (control: 65.2±7.0 pA; AP5: 39.3±5.1 pA; n=10, p<0.001) (Fig. 2D). In a subset of experiments, AP5 inhibition of IAMPA was tested in the absence of picrotoxin (Fig. 2C, inset). Pretreatment with AP5 efficiently reduced IAMPA in the absence of picrotoxin at Vholding of –20 mV in normal aCSF (control: 55.8±6.7 pA; AP5: 35.3±5.3 pA; n=4, p<0.01).
These results suggested that NMDAR activation enhanced the steady-state activation of AMPARs, altering the neuronal response to the receptor agonists in SON MNCs.
Roles of eNMDAR in IAMPA potentiation in SON MNCS
In the next experiments, we investigated whether eNMDARs that generate tonic INMDA contribute to IAMPA facilitation in SON MNCs. To isolate eNMDAR function, we adopted an eNMDAR selective antagonist, memantine [2122]. To investigate the role of eNMDARs in IAMPA facilitation, we recorded and compared IAMPA amplitude in the absence and presence of memantine in low-Mg2+ aCSF (Fig. 3). Memantine induced an outward shift in Iholding (13.2±3.8 pA, n=6), which is comparable to tonic INMDA uncovered by AP5 (12.1±2.5 pA, n=7, p>0.7) in low-Mg2+ aCSF.
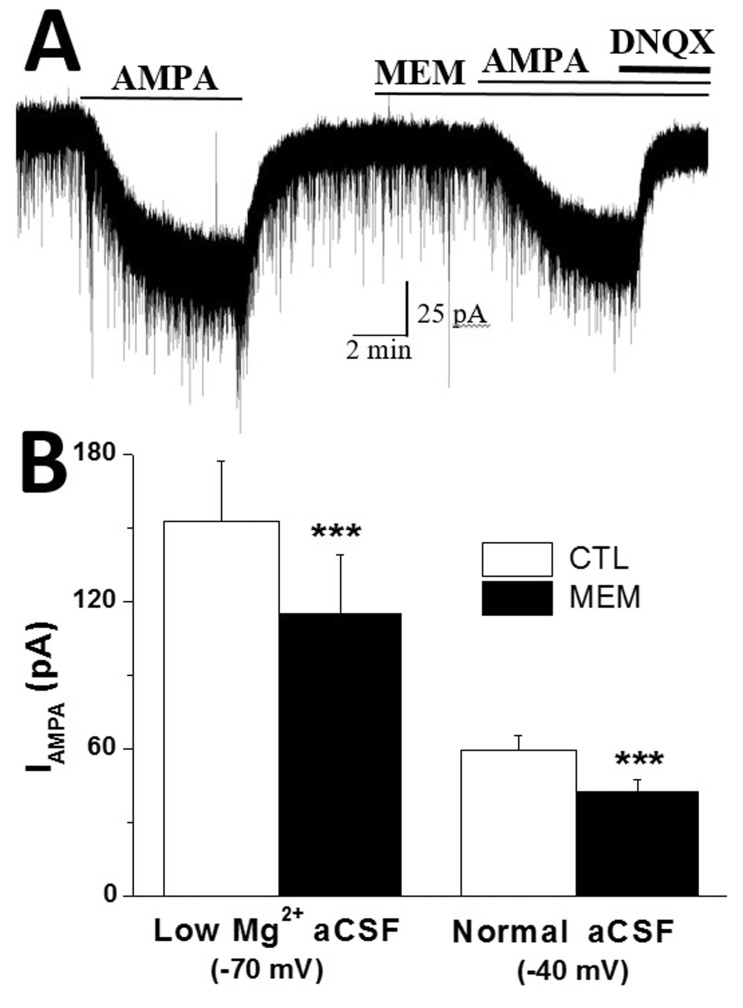 | Fig. 3Extrasynaptic NMDARs mediate IAMPA facilitation in SON MNCs.(A) Representative current traces showing that the extrasynaptic NMDAR antagonist, memantine (MEM), attenuated IAMPA in low-Mg2+ aCSF (Vholding –70 mV). (B) Effects of memantine on IAMPA amplitude in low-Mg2+ aCSF (Vholding –70 mV, n = 8) and normal aCSF (Vholding –40 mV, n = 6) are summarized. Effects of MEM on the first and second IAMPA were pooled as in Fig. 2. ***p<0.001 compared to respective control.
|
Pretreatment with memantine reversibly and significantly reduced IAMPA amplitudes from 152.3±24.4 pA to 114.8±24.0 pA (n=8) in low-Mg2+ aCSF (p<0.05) (Fig. 3A and 3B), while it failed to affect IAMPA in normal aCSF (control: 108.8±12.7 pA; memantine: 105.0±13.2 pA; n=6). The inhibition rate of IAMPA by pretreatment with memantine (65.8±5.4% of control, n=8) was comparable to that of AP5 (60.3±4.06% of control, n=12, p>0.4) in low-Mg2+ aCSF. Pretreatment with memantine also significantly reduced IAMPA at Vholding of –40 mV in normal aCSF (control: 59.2±6.1 pA; memantine: 42.3±5.1 pA; n=6, p<0.01) (Fig. 3B).
These results suggested that eNMDAR activation facilitates IAMPA in SON MNCs.
IAMPA potentiation by NMDARs during chronic dehydration
To investigate the functional significance of IAMPA facilitation by tonic INMDA in SON MNCs, we examined and compared the IAMPA potentiation by INMDA in euhydrated (EU) and chronic dehydrated (DE) rats (Fig. 4). Chronic dehydration with a 7-day salt loading (2% NaCl) protocol increased plasma osmolarity from 306.7±2.3 mOsm (n=4) to 378.5±7.1 mOsm (n=5). Consistent with a previous report [23], DE significantly increased EPSC frequency in SON MNCs (EU: 1.81±0.35 Hz, n=6; DE: 4.09±0.62 Hz, n=7).
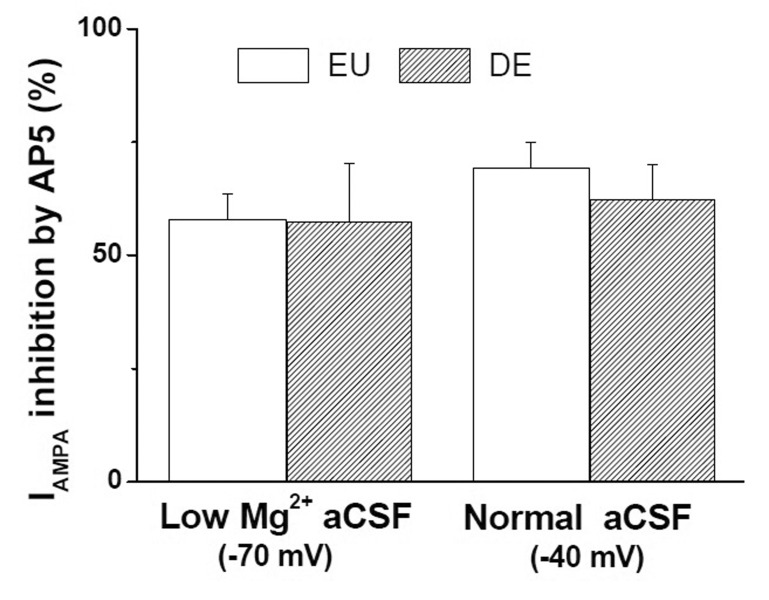
Bath application of AMPA induced reproducible IAMPA in DE SON MNCs as in EU SON MNCs. Pretreatment with AP5 significantly reduced IAMPA in low-Mg2+ aCSF at Vholding of –70 mV (control: 68.6±15.0; AP5: 45.9±12.8 pA; n=8, p<0.05) and in normal aCSF at Vholding of –40 mV (control: 70.1±10.4 Pa; AP5: 47.9±8.6 pA; n=8, p< 0.001) in DE SON MNCs. Although IAMPA amplitudes were slightly lower in DE than in EU neurons, the difference did not reach statistical significance (p>0.2 in both cases). Furthermore, pretreatment with AP5 inhibited IAMPA at similar rates in EU and DE SON MNCs (Fig 4), suggesting that IAMPA potentiation by INMDA was preserved in DE SON MNCs.
Go to :
DISCUSSION
The main findings of this study may be summarized as follows: 1) glutamate induced the tonic activation of AMPARs in a tonic INMDA-dependent manner; 2) tonic INMDA mediated by eNMDARs facilitated AMPA-induced steady sate inward currents (IAMPA); and 3) IAMPA potentiation by eNMDARs was preserved in SON MNCs during chronic dehydration. To our knowledge, these data are the first to demonstrate that eNMDAR activity modulates the steady-state sensitivity of the AMPA receptor channels to their agonists in neuroendocrine systems.
Tonic INMDA mediated by eNMDARs in SON MNCs
Although receptors are often considered extrasynaptic if they are located more than 100 nm from the postsynaptic density, the precise delineation of the synaptic-extrasynaptic receptors seems to be specific to the parameter under considerations [24]. In terms of electrophysiological activities, synaptic NMDARs are defined as receptors recruited in response to spontaneous glutamate release, generating EPSCs, or during low-frequency afferent stimulation (less than 0.05 Hz), while eNMDARs correspond to those not activated during such conditions. In this sense, there is a general consensus that eNMDARs are responsible for the persistent tonic excitatory current in SON MNCs [625], while their synaptic counterparts mediate conventional EPSCs. In the present study, Iholding shift by a NMDAR antagonist, AP5, supported the presence of a persistent inward current with basal glutamate release mediated by NMDARs in SON MNCs. Combined with the fact that memantine failed to affect the basic properties of glutamate EPSCs in SON MNCs [6], our results showed that memantine, a selective eNMDAR blocker, mimicked the AP5-induced Iholding shift, which confirmed the notion that eNMDARs generate tonic INMDA in the neurons. The subunit composition of the eNMDAR mediating tonic INMDA in various brain regions has not been well clarified, while eNMDARs containing the NR2B subunit have been known to be partly responsible for tonic INMDA [6]. Future studies are warranted to delineate the subunit composition of the eNMDARs mediating IAMPA facilitation in SON MNCs.
Modulation of AMPARs function by tonic INMDA in SON MNCs
Our results showing that IAMPA was inhibited by AP5 or extracellular Mg2+ in low-Mg2+ aCSF suggested that activated NMDARs facilitated AMPARs function in SON MNCs. Furthermore, similar inhibitory effects of AP5 and memantine on IAMPA facilitation suggested that eNMDARs facilitated AMPAR function in the neurons. Combined with the fact that IGLU amplitudes were dependent on AP5 (Fig. 1), these results suggested that eNMDAR activity modulates the steady-state sensitivity of AMPA receptors to their agonists in SON MNCs. Given that binding glutamate/AMPA to the AMPA receptor results in Na+ influx, which causes depolarization of the membrane, our results showing that AP5 inhibited IAMPA/IGLU could indicate that AP5 inhibited eNMDAR activated by the agonists. However, our results showing that AMPARs antagonists completely abolished IAMPA in both the absence and presence of NMDAR antagonists (Fig. 2) argued against this possibility. It is not likely that a bath application of AMPA recruited additional NMDARs generating INMDA in our recording conditions. This idea was further supported by the finding that IAMPA was insensitive to the following application of AP5 (data not shown).
In general, there has been a consensus that extrasynaptic glutamate receptors generate slow, persistent tonic currents, while their synaptic counterparts mediate AMPAR- or NMDAR-EPSCs. In the present study, it is not clear whether extrasynaptic AMPARs (eAMPARs) generate IAMPA, and, if so, what portion of the currents is mediated by eAMPARs in SON MNCs. However, it is interesting to note that eAMPARs are highly mobile and move rapidly between the plasma membrane and the intracellular compartments by exocytosis and endocytosis, and diffuse laterally to and from synaptic sites [26272829]. Such continuous AMPAR exchanges between synapses and different cellular pools ensures a dynamic fit of synaptic AMPAR numbers in synaptic plasticity, including long-term potentiation (LTP). A prevailing two-step model is that NMDAR-dependent LTP is mediated by surface insertion and synaptic delivery of AMPARs, in which AMPARs deliver to the extrasynaptic sites first, and their synaptic targeting requires synaptic NMDAR activation that likely triggers the signal transduction cascade necessary to anchor AMPARs in the synapse. Given that activated eNMDARs cause Ca2+ influx [9], it is reasonable to assume that eNMDARs regulated AMPARs trafficking in the present study, resulting in IAMPA facilitation. As a gliotransmitter, ATP has been known to activate purinergic receptors in PVN MNCs, promoting the insertion of AMPARs at the surface and, strengthening the excitatory synapses [14]. However, it is noteworthy that NMDAR-induced CaM-KII activation alters AMPARs channel properties in both a Ca2+-dependent and a Ca2+-independent manner [3031]. It is also possible that INMDA facilitated AMPAR-mediate currents via increased single channel conductance and/or open channel probability of AMPAR activated channels in SON MNCs. Future studies are warranted to delineate the cellular mechanisms of eNMDAR-mediated IAMPA facilitation in SON MNCs.
Functional significance of IAMPA potentiation by eNMDARs in SON MNCs
A peculiar property of the SON is that it undergoes anatomical remodeling under certain physiological conditions, such as lactation and chronic dehydration. This remodeling includes a reduction in astrocytic coverage of neurons [32] and is associated with increased extracellular levels of glutamate in the SON [33]. The increased extracellular level of glutamate in the nucleus is also in agreement with enhanced EPSC frequency [23] and increased numbers of excitatory synaptic contacts in SON MNCs. Combined with our results showing that IAMPA facilitation by INMDA is conserved in DE SON MNCs, these results suggested that IAMPA facilitation by INMDA could contribute to enhanced neuronal activity and hormone release from the neurons during the osmotic challenge.
Spontaneous glutamate release from astrocytes synchronizes neuronal activity via eNMDAR activation [3435], and increases the frequency of AMPAR-mediated EPSCs via the activation of metabotropic glutamate receptors facilitating presynaptic release in CA1 pyramidal neurons [36] and kainite receptors in hippocampal interneurons [37], respectively. Combined with the fact that Ca2+ influx triggers the signal transduction cascade necessary for anchoring AMPARs in synapses [2638], our results showing that eNMDAR activation potentiated AMPAR-mediated steady-state currents in SON MNCs are in line with astrocytic glutamate release altering AMPARs, resulting in strengthening of excitatory synapses in SON MNCs.
In summary, our results support the view that tonic INMDA generated by activated eNMDARs facilitates AMPA receptor function in SON MNCs, which gives the gliotransmitter glutamate the ability to regulate postsynaptic efficacy during normal and physiological challenges, including chronic dehydration.
Go to :
 XML Download
XML Download