

INTRODUCTION
The polyphenol resveratrol is enriched in the skins of red grapes [12]. It has attracted increasing scientific attention due to its cardiovascular benefits and potent antitumor activity [34]. In obese rodents, resveratrol treatment produces various health benefits, including enhanced vascular function, decreased restenosis and hypertension, reduced inflammation and a gene expression pattern resembling that which occurs during caloric restriction [567]. Despite the significance of resveratrol for protecting against cardiovascular diseases, whether and how resveratrol produces these beneficial effects are still poorly understood.
The blood-brain barrier (BBB) protects the brain from potentially neurotoxic substances and facilitates the exchange of nutrients and waste products between the brain and the blood, thus maintaining an optimal extracellular environment for neuronal function [8]. A major role of the BBB is strict regulation of paracellular permeability. This is primarily mediated by the tight junctions between endothelial cells of the capillary, which restrict paracellular movement of solutes, ions, and water. Tight junctions are dynamic and highly regulated structures, and decreased tight-junction protein expression or variations in subcellular localization are associated with alterations in paracellular permeability [910].
The AMP-activated protein kinase (AMPK) is a heterotrimeric protein composed of α, β, and γ subunits [11]. The β subunit imparts catalytic activity, while the γ subunit contains a glycogen-binding domain that also regulates the activity and the γ subunit forms the broad base of the protein and is required for AMP binding. AMPK is well-conserved among eukaryotic cells [1213]. Previous studies indicated that AMPK can be activated by resveratrol [14] and its activation counteracts oxidative stress by suppressing NAD(P)H oxidase-derived reactive oxygen species (ROS) production in endothelial cells [1516]. At present, the specific signaling pathways within brain endothelial cells that target disengagement of the tight junction remain elusive. We hypothesized that AMPK activation via suppression of ROS contributes to the maintenance of the BBB integrity. Here we provide evidences that LPS regulates tight-junction dynamics in brain endothelial cells via NAD(P)H oxidase-derived ROS and show that AMPK activation by protects tight-junction proteins, and consequently BBB integrity, by suppressing the expression of NAD(P)H oxidase.
Go to : 
METHODS
Reagents
Human brain microvascular endothelial cells (HBMECs), cell culture media, and growth supplements were obtained from Sciencell (Carlsbad, CA, USA). Primary antibodies against phospho-AMPKα (Thr-172), AMPKα, GAPDH, and all secondary antibodies were obtained from Cell Signaling Technology (Danvers, MA, USA). Antibodies against occludin and anti-claudin-5 were from Invitrogen Life Technologies (Carlsbad, CA, USA). Antibodies against the NAD(P)H oxidase subunits (p47phox, p67phox) were from Santa Cruz Biotechnology (Santa Cruz, CA, USA). FITC-dextran was from Sigma–Aldrich (St. Louis, MO, USA). A stock solution of LPS (Escherichia coli O111:B4; Calbiochem, Germany) was prepared in PBS. The concentration of LPS was selected to mimic sepsis [17].
Cell culture
HBMECs were cultured in endothelial cell medium mixed with astrocyte-conditioned medium (1:1). All cells were incubated at 37℃ in a humidified atmosphere of 5% CO2 and 95% air. Confluent cells were maintained in 1% fetal calf serum and exposed to LPS after overnight starvation. In all experiments, cells were used between passages 3 and 8.
Adenovirus infection
HBMECs were infected with adenoviruses encoding Cu-Zn-SOD, green fluorescent protein (GFP), or constitutively active AMPK (AMPK-CA) (multiplicity of infection 50) as described previously [16]. Under these conditions, the infection efficiency was typically >80% as determined by green fluorescent protein expression.
Measurement of ROS
Intracellular ROS was measured using the dihydroethidium fluorescence/HPLC assay with minor modifications [18]. Briefly, before the end of experiment, HBMECs were incubated with 0.5 µM dihydroethidium for 30 min. After incubation, the cells were harvested and extracted with methanol. Oxyethidium (a product of dihydroethidium and superoxide anions) and ethidium (a product of dihydroethidium autoxidation) were separated and quantified by C-18 HPLC column (mobile phase: gradient of acetonitrile and 0.1% trifluoroacetic acid) coupled with a fluorescence detector. Superoxide anion production was determined by the conversion of dihydroethidium into oxyethidium.
Permeability of the BBB in vitro
HBMECs were cultured on Costar Transwell filters (pore size 0.4 µm; Corning, NY, USA) that were coated on the upper side with fibronectin (Sigma–Aldrich) in endothelial cell growth medium mixed with astrocyte-conditioned medium at a 1:1 ratio as described previously [19]. Permeability for FITC-dextran (150 kDa) was assayed as described previously. At various time points after LPS treatment, samples were collected from the acceptor chambers for measurement of fluorescence intensity at excitation 485 nm and emission 520 nm using a microplate reader (Synergy HT; BioTek Instruments, Winooski, VT, USA).
Determination of trans-endothelial electrical resistance
Determination of trans-endothelial electrical resistance across the monolayer of HBMECs was measured using the Millicell-ERS (Millipore). Briefly, 250 µl of cell suspension (8×105 cells/ml) was seeded in each collagen-coated well. When confluent monolayer reached maximum resistance (150 Ω/cm2), the endothelial integrity of HBMECs treated with or without LPS was measured in real time as described previously [19]. Values are presented as Ω/cm2 of culture well. The trans-endothelial electrical resistance of cell-free wells was subtracted from the obtained values.
Western blotting
Cells were washed with phosphate-buffered saline (PBS) and homogenized in RIPA lysis buffer (Cell Signaling Technology). Cell lysates were sonicated twice for 10 s in an ultrasonic dismembrator with 10% output (Model 500; Fisher Scientific) and centrifuged at 14,000 g for 20 min at 4℃. The supernatants were subjected to Western blot analysis, as described previously. GAPDH was used as an internal control.
Measurement of NAD(P)H oxidase activity
NAD(P)H oxidase activity was measured as described pre viously [19]. Briefly, 20 µg protein was incubated with dihydroethidium (10 µM) and DNA (1.25 µg/ml) in PBS with the addition of NAD(P)H (50 µM), in a final volume of 120 µl. Incubations were performed for 30 min at 37℃ in the dark. Fluorescence intensity was recorded in a microplate reader (excitation 490 nm and emission 590 nm).
Statistical analysis
Data are expressed as means±SEM of the mean unless otherwise noted. Statistical comparison was performed using a one-way analysis of variance followed the Student-Newman-Keuls test using SPSS 10.0 software. Differences were considered significant at p<0.05.
Go to :
RESULTS
LPS disrupted BBB integrity
We first determined the effect of LPS on the paracellular permeability in cultured HBMEC monolayer on Transwell filters. As depicted in Fig. 1A, 1 µg/ml LPS significantly increased the leakage of FITC-dextran of 150 kDa from 8 hours and reached the maximal effect at 24 hours.
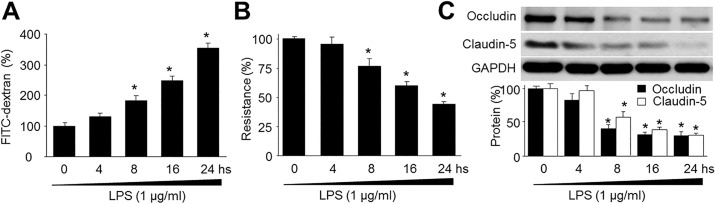 | Fig. 1LPS decreases blood-brain barrier integrity and reduces the expression of tight-junction proteins in human brain microvascular endothelial cells (HBMECs).(A) Confluent monolayer of HBMECs was cultured on Transwell filters and the diffusion of FITC-conjugated dextran (150 kDa; 100 µg/ml) in LPS (1 µg/ml) was measured at various time points. Control was set up as 100%. Data are expressed as the mean±SEM. N is 5 in each group. *p<0.05 vs control. (B) Trans-endothelial electrical resistance was measured in real time in confluent monolayer of HBMECs. Control was set up as 100%. Data are expressed as the mean±SEM. N is 5 in each group. *p<0.05 vs control. (C) Representative Western blot of tight-junction proteins (occludin and claudin-5). The blot is representative of three blots from three independent experiments. *p<0.05 vs control.
|
To investigate the influence of LPS on the integrity of the BBB in vitro, trans-endothelial electrical resistance of confluent human brain endothelial monolayer was measured at various time points after LPS exposure. As shown in Fig. 1B, LPS reduced trans-endothelial electrical resistance across a monolayer of human brain microvascular endothelial cells in a time-dependent manner.
LPS reduced expressions of tight-junction proteins
BBB integrity is mainly dependent on the presence of tight junctions [20]. Therefore, LPS-induced alterations in tight junctions may be responsible for the increased BBB permeability. To determine the effects of LPS on tight junctions in brain endothelium, we measured real-time dynamics of the expression of the tight-junction proteins, such as occludin and claudin-5, which are essential for tight-junction maintenance. As shown in Fig. 1C, expression levels of claudin-5 and occludin were significantly decreased by LPS from 8 hours treatment. These results suggest that BBB leakage under sepsis condition was due to the loss of tight-junction proteins.
LPS increased production of superoxide anions
Evidences have indicated that ROS disrupts endothelial cell tight junctions, resulting in increased paracellular permeability [21]. To determine whether ROS caused the tight-junction alterations induced by LPS, we next assayed superoxide anion levels in HBMECs after exposure to LPS. Exposure to LPS time-dependently caused a greater production of superoxide anions in HBMECs (Fig. 2A).
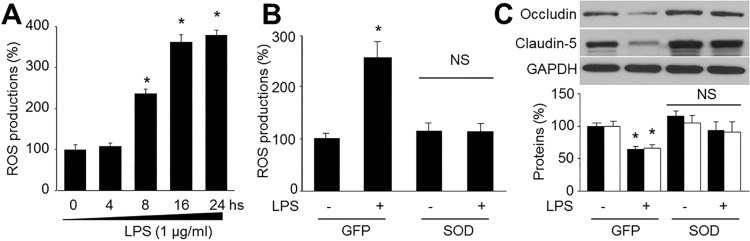 | Fig. 2Reactive oxygen species (ROS) mediates LPS-reduced tight-junction protein expressions in human brain microvascular endothelial cells (HBMECs).(A) Confluent monolayer of HBMECs were cultured in LPS (1 µg/ml) was measured at various time points. ROS productions were assayed by DHE fluorescence. Data are expressed as the mean±SEM. N is 5 in each group. *p<0.05 vs control. (B and C) Cultured HBMECs were infected with adenovirus containing superoxide dismutase (SOD) for 48 hours and then exposure to LPS of 1 µg/ml for 24 hours. (B) Superoxide anion productions and (C) levels of occludin and claudin-5 expression were measured. N is 5 in each group. *p<0.05 vs control.
|
Overexpression of SOD prevented LPS-induced reduction of tight-junction proteins
To further confirm whether the loss of tight-junction proteins was induced by ROS, HBMECs were infected with adenoviruses encoding Cu-Zn-SOD, a major antioxidant enzyme. In Fig. 2B and 2C, overexpression of Cu-Zn-SOD abolished the reduction of occludin and claudin-5 expressions in LPS-treated HBMECs, as well as increased ROS production. Taken together, these results suggest that the suppression of tight-junction proteins by LPS is mediated by superoxide anions.
Inhibition of NAD(P)H oxidase attenuated LPS-induced reduction of tight-junction proteins
To study the role of NAD(P)H oxidase in ROS-induced tight-junction alterations, we measured NAD(P)H oxidase activity in HBMECs after exposure to LPS. As shown in Fig. 3A, the activity of NAD(P)H oxidase was markedly increased by LPS. Apocynin, a potent pharmacologic inhibitor of NAD(P)H oxidase, significantly reduced the LPS-induced increase in ROS productions (Fig. 3B).
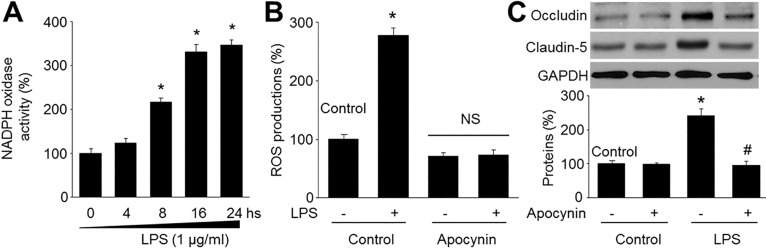 | Fig. 3LPS decreases tight-junction protein levels via upregulation of NAD(P)H oxidase in microvascular endothelial cells.(A) Confluent monolayer of HBMECs were cultured in LPS (1 µg/ml) was measured at various time points. NAD(P)H oxidase activity was was assayed by DHE fluorescence. Data are expressed as the mean±SEM. N is 5 in each group. *p<0.05 vs control. (B and C) Cultured HBMECs were pretreated with apocynin (10 µM) for 30 mins and then co-incubated with LPS (1 µg/ml) for 24 hours. (B) ROS productions were detected by DHE. N is 5 in each group. *p<0.05 vs control. NS indicates no significance. (C) Levels of occludin and claudin-5 expression were measured by Western blot. The blot is representative of three blots from three independent experiments. *p<0.05 vs control. #p<0.05 vs LPS alone.
|
We further determined whether pharmacological inhibition of NAD(P)H oxidase affected the levels of tight-junction proteins. As expected, apocynin significantly attenuated the reduction in occludin and claudin-5 caused by LPS (Fig. 3C). Taken together, these results suggest that LPS reduces the level of tight-junction proteins via NAD(P)H oxidase.
AMPK activation by resveratrol inhibited LPS-induced oxidative stress in HBMECs
AMPK is reported to play a key role in maintaining redox homeostasis in endothelial cells, and inhibition of AMPK causes aberrant oxidative stress [2223]. We therefore examined whether alteration of AMPK activity affected ROS productions after exposure of HBMECs to LPS. As expected, resveratrol, which is a known AMPK activator [24], significantly increased AMPK and ACC phosphorylations (Fig. 4A), consistent with other reports [2526]. Importantly, AMPK activation by resveratrol attenuated the LPS-induced elevation of ROS productions (Fig. 4B). Consistently, adenoviral overexpression of AMPK-CA mutants, but not the GFP control, significantly decreased the induction of superoxide anions by LPS (Fig. 4C).
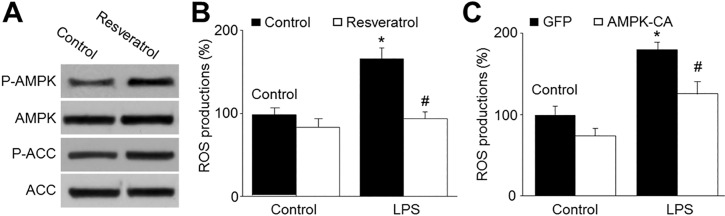 | Fig. 4AMPK activation by resveratrol reduces LPS-enhanced production of reactive oxygen species in HBMECs.(A) Cultured HBMECs were treated with resveratrol (10 µM) for 24 hours. The levels of pAMPK and pACC were measured by Western blotting. The picture is a representative picture from 3 independent experiments. (B) Cultured HBMECs were pretreated with resveratrol (10 µM) for 30 min and then co-incubated with LPS (1 µg/ml) for 24 hours. ROS productions were measured by DHE/HPLC. Data are expressed as the mean±SEM. N is 5 in each group. *p<0.05 vs control. #p<0.05 vs LPS alone. (C) Cultured HBMECs were infected with adenovirus containing constitutively active AMPK (AMPK-CA) mutant for 48 hours and then incubated with LPS (1 µg/ml) for 24 hours. ROS productions were measured by DHE/HPLC. Data are expressed as the mean±SEM. N is 5 in each group. *p<0.05 vs GFP alone. #p<0.05 vs LPS alone.
|
Resveratrol via AMPK activation prevented LPS-induced reduction in tight-junction proteins
Next, we examined the effects of resveratrol on tight-junction proteins. To determine whether AMPK activation by resveratrol can prevent loss of tight-junction proteins, we tested the effects of resveratrol on the expression of occludin and claudin-5 after LPS exposure. As expected, resveratrol significantly alleviated the LPS-induced loss of tight-junction proteins (Fig. 5A), and improved the morphology of tight junction as determined by staining F-actin (Fig. 5B).
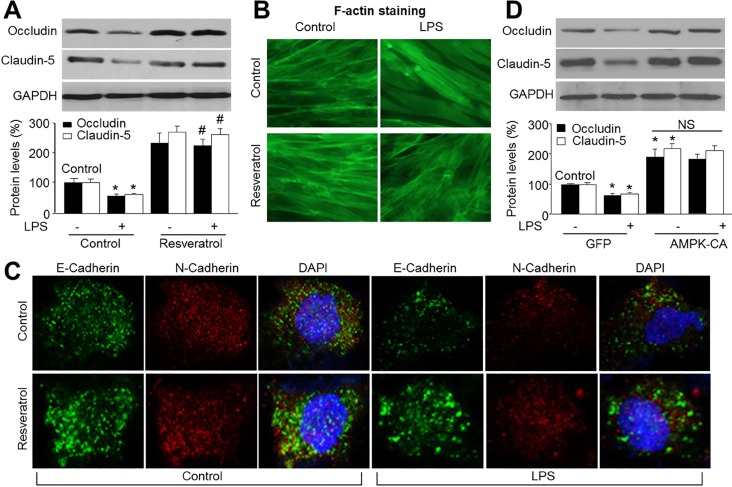 | Fig. 5AMPK activation by resveratrol prevents LPS-induced loss of tight-junction proteins.(A~C) Cultured HBMECs were pretreated with resveratrol (10 µM) for 30 min and then co-incubated with LPS (1 µg/ml) for 24 hours. (A) Expressions of tight junction protein, occludin and claudin-5, were measured by Western blot. Data are expressed as the mean±SEM. N is 3 in each group. *p<0.05 vs control. #p<0.05 vs LPS alone. NS indicates no significance. (B)The morphology of tight-junction was determined by using staining F-actin. (C) Double-labeled immunofluorescence analysis of E-Cadherin (green) and N-Cadherin (red) proteins. Blue, DAPI stained nucleus. (D) Cultured HBMECs were infected with adenovirus containing constitutively active AMPK (AMPK-CA) mutant for 48 hours and then incubated with LPS (1 µg/ml) for 24 hours. Expressions of tight junction protein, occludin and claudin-5, were measured by Western blot. Data are expressed as the mean±SEM. N is 3 in each group. *p<0.05 vs GFP alone. NS indicates no significance.
|
The cadherin family, E-cadherin and N-cadherin, share many structural and functional features, which establish Ca2+-dependent homophilic cell-cell adhesion with their extracellular domains and connect with catenin at their intracellular domains [27]. In this study, the effects of resveratrol on expression and distribution of E-cadherin and N-cadherin were examined by immunofluorescence. As shown in Fig. 5C, resveratrol could markedly increase the expression of E-cadherin, but did not affect N-cadherin expression. There are no obvious changes of localization could be found.
To exclude potential off-target effects of resveratrol, we also investigated the effects of AMPK on tight-junction proteins using genetic approaches. Adenoviral overexpression of AMPK-CA mutants accentuated the reduction in tight-junction proteins caused by LPS (Fig. 5D).
AMPK activation prevented LPS-induced upregulation of NAD(P)H oxidase
NAD(P)H oxidase consists of membrane-bound gp91phox and p22phox as well as cytosolic subunits such as p47phox, p67phox, and small GTPase Rac1 [162829]. As AMPK activation suppressed LPS-induced ROS productions and the loss of tight junctions in HBMECs, we next determined whether these effects of AMPK were mediated by decreasing NAD(P)H oxidase. As expected, activation of AMPK with resveratrol significantly alleviated the increased expressions of the essential subunits of p67phox and p47phox caused by LPS in HBMECs (Fig. 6A). Consistently, overexpression of AMPK-CA prevented LPS-induced upregulations of p67phox and p47phox (Fig. 6B). Taken together, these data suggest that AMPK activation maintains the levels of tight-junction proteins in HBMECs by inhibiting LPS-enhanced NAD(P)H oxidase expression.
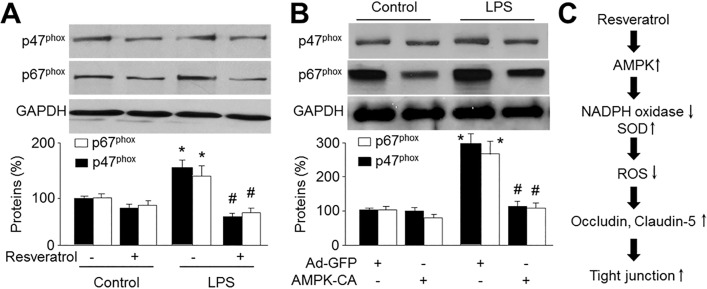 | Fig. 6AMPK activation prevents LPS-induced upregulation of NAD(P)H oxidase subunits.(A) Cultured HBMECs were pretreated with resveratrol (10 µM) for 30 min and then co-incubated with LPS (1 µg/ml) for 24 hours. Expressions of NAD(P)H oxidase subunits, p47phox and p67phox, were measured by Western blot. Data are expressed as the mean±SEM. N is 3 in each group. *p<0.05 vs control. #p<0.05 vs LPS alone. NS indicates no significance. (B) Cultured HBMECs were infected with adenovirus containing constitutively active AMPK (AMPK-CA) mutant for 48 hours and then incubated with LPS (1 µg/ml) for 24 hours. Expressions of NAD(P)H oxidase subunits, p47phox and p67phox, were measured by Western blot. Data are expressed as the mean±SEM. N is 3 in each group. *p<0.05 vs GFP alone. NS indicates no significance. (C) Proposed molecular mechanism for protective effect of AMPK activation on BBB integrity.
|
Go to :
DISCUSSION
The most important findings of this study is that LPS reduces the levels of tight-junction proteins. Additionally, we found that LPS significantly increased levels of ROS and that adenoviral overexpression of SOD significantly attenuated the LPS-induced suppression of tight-junction proteins. Thus, it is likely that the LPS-induced reduction in tight-junction proteins is mediated by endogenous ROS. Our observations are consistent with previous studies indicating that oxidative stress disrupts endothelial tight junctions, resulting in increased paracellular permeability [30]. In this study, we also found that administration of LPS to monolayer of HBMECs induced a time-dependent decrease in the trans-endothelial electrical resistance, accompanied by an enhanced permeability of the monolayer to the hydrophilic model compound FITC-dextran. Tight-junction proteins are the main determinants of low permeability and high electrical resistance in vitro. Consistently, our studies revealed that treatment of HBMEC monolayer with LPS resulted in a time-dependent decrease in tight-junction protein expression, which paralleled the decreased trans-endothelial electrical resistance and the increased amount of FITC-dextran that crossed the monolayer. Indeed, published studies [2829] have shown that diseases of the periphery, such as diabetes, have deleterious effects on the BBB that may contribute to the increased risk of cerebrovascular disease and neurodegeneration associated with these conditions.
Another important finding of this study is that resveratrol prevents LPS-induced BBB dysfunction. AMPK is known to play a key role in endothelial cell biology. In this study, we found that AMPK activity declined upon long-term exposure to LPS. Activation of AMPK by resveratrol suppressed ROS productions and reversed the tight-junction protein levels. Moreover, we have identified NAD(P)H oxidase as the main source of ROS induced by LPS. Our data showed that expression of NAD(P)H oxidase subunits was unregulated in the presence of LPS. Accordingly, inhibition of NAD(P)H oxidase abolished ROS productions and consequently significantly reversed the level of tight-junction proteins. Importantly, resveratrol markedly decreased the unregulated expression of NAD(P)H oxidase subunits, suggesting that the effects of resveratrol on tight-junction protein levels are directly mediated by inhibition of NAD(P)H oxidase-derived ROS. Indeed, recent studies suggest that AMPK activation improves endothelial function by counteracting oxidative stress in endothelial cells [31].
In summary, our results suggest that LPS disrupts the expression of tight-junction proteins in the BBB via production of ROS and that activation of AMPK by resveratrol helps to maintain the integrity of the BBB by suppression of NAD(P) H oxidase (Fig. 6C). Thus, AMPK might be important in maintaining the integrity of the BBB in patients with sepsis and AMPK inhibition might play an important role in the initiation and progression of neurological disorders in diabetes.
Go to :