

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Like Panax ginseng, Cordyceps is a genus of insect-parasitizing fungus that is used as a traditional herbal medicine in Korea and China [123]. Treatment with Cordyceps species relieves various immunological disorders including respiratory, pulmonary, and cardiovascular diseases [23]. Systemic studies have found that Cordyceps species have numerous biological activities including anti-metastatic, anti-oxidative, anti-inf lammatory, antimicrobial, anti-aging, insecticidal, hypolipidemic, hypoglycemic, and neuroprotective effects. Furthermore, Cordyceps has been considered a promising drug for treating cancer due to its antiproliferative effect [1234].
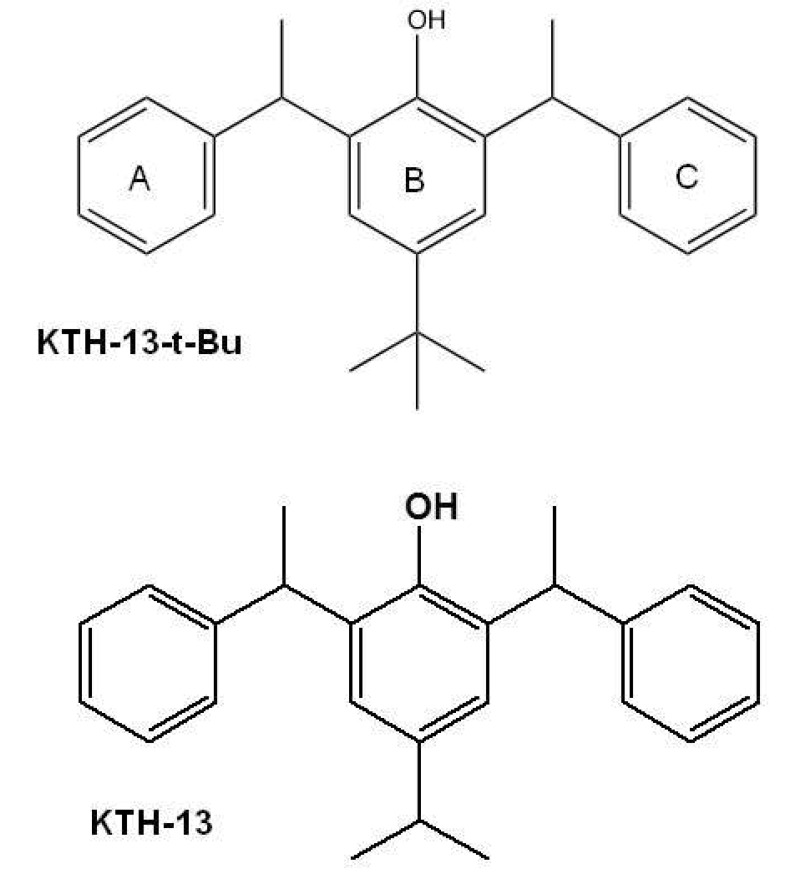
Cordyceps bassiana is a Cordyceps species that can be used as an effective herbal remedy. The butanol fraction of C. bassiana (Cb-BF) has been reported to inhibit interleukin (IL)-12 and tumor necrosis factor (TNF)-α by suppression of inflammatory signaling cascades including spleen tyrosine kinase (Syk), janus kinase (JAK)-2, and extracellular signal-regulated kinase (ERK) [5]. In addition, Cb-BF was revealed to diminish the proliferation of splenic lymphocytes, and to ameliorate various symptoms in atopic dermatitis [6]. From the same fraction, we also isolated an anti-cancer inhibitory compound with pro-apoptotic activity, KTH-13 [4-isopropyl-2,6-bis(1-phenylethyl)phenol] (Fig. 1), as well as other non-active compounds such as (E)-2-(2-(3-acetoxy-2-(acetoxymethyl)propyl)-5-(((2-hydroxyethoxy)methyl)amino)-5-oxopent-3-en-1-yl)propane-1,3-diyl diacetate (Bassiamide A), (E)-2-(15-(3-acetoxy-2-(acetoxymethyl)propyl)-5-(acetoxymethyl)-2,12-dioxo-3,6,8,11-tetraoxahexadec-13-en-16-yl)propane-1,3-diyl diacetate (Bassiamate), 1-(N-methylbenzamido)-3-(tetradecanoyloxy)propan-2-yl benzoate, and (Z)-1-acetoxy-3-(oleoyloxy)propan-2-yl 4-isopropylcyclohex-2-enecarboxylate [7]. To improve the anti-proliferative activity and production yield, we employed a total synthetic approach and further derivatized KTH-13 to obtain chemical analogs. By this process, we synthesized an analog of KTH-13, KTH-13-t-Bu [4-(tert-butyl)-2,6-bis(1-phenylethyl)phenol (Fig. 1)], as a new synthetic compound. In this study, we examined its anti-cancer activity to demonstrate the biological effect of this compound in terms of inhibiting proliferation and promoting apoptosis in various cancer cells.
METHODS
Materials
KTH-13-t-Bu was supplied by Prof Lee, Yunmi (Kwangwoon University, Seoul). This compound was more than 98% pure according to HPLC analysis. (3-4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide, tetrazole (MTT) and bisbenzimide were purchased from Sigma Chemical Co. (St. Louis, MO, USA). Fetal bovine serum (FBS), Dulbecco's modified Eagle's medium (DMEM) and penicillin/streptomycin were obtained from Thermo Fisher Scientific Inc. (Waltham, MA, USA). C6 glioma, MDA-MB-231, LoVo, and HCT-15 cells were purchased from ATCC (Rockville, MD, USA). The FITC annexin V Apoptosis Detection Kit I was purchased from BD biosciences (San Diego, CA, USA). Antibodies against total caspases (3, 8, and 9) and phospho- and total forms of Bax, Bcl-2, STAT3, PI3K/p85, Src, and β-actin were obtained from Cell Signaling (Beverly, MA, USA).
Cell culture
C6 glioma, MDA-MB-231, LoVo, and HCT-15 cells were cultured in DMEM with 5% heat-inactivated FBS and 1% penicillin/streptomycin at 37℃ in 5% CO2, as reported previously [8]. For each experiment, cells were detached with trypsin/EDTA solution. The cell density used in our experiments is 5×105 cells/ml.
Cell viability test
After pre-incubating C6 glioma, MDA-MB-231, LoVo, and HCT-15 cells (5×105 cells/ml) for 18 h, KTH-13-t-Bu was added to the cells and incubated for 6 or 24 h with 5% FBS. The effect of this compound on cell proliferation was then evaluated by using a conventional MTT assay [910]. Ten microliters of MTT solution (10 mg/ml in phosphate-buffered saline (PBS), pH 7.4) was added to the cultures, and the cells were cultured for 3 h. The incubation was stopped by adding 15% sodium dodecyl sulphate to each well to solubilize the formazan [11]. The absorbance at 570 nm (OD570~630) was assessed by using a Spectramax 250 microplate reader.
Morphological change test
C6 glioma cells were incubated with KTH-13-t-Bu (0 to 50 µM) for the indicated times. Images of the cells in culture at each time point were obtained by using an inverted phase contrast microscope, attached to a video camera with NIH image software [12].
FITC annexin V-PI staining apoptosis assay
Apoptosis was determined with a FITC annexin V Apoptosis Detection Kit I that detects phosphatidylserine-based changes in cell membranes [1314]. Cells were plated in 12-well culture plate at a seeding density of 4×105 cells/dish (5×105 cells/ml), and KTH-13-t-Bu was added to the culture medium to the specified concentration. Vehicle alone was used as the untreated control. The subsequent procedures were conducted according to the instructions provided by the manufacturer. Briefly, at each time point, cells were harvested, washed twice with PBS and resuspended in 1X binding buffer. Annexin-V FITC and PI were added and incubated for 15 min at room temperature (25℃) in the dark. Fluorescence from 1×105 cells were detected with a BD FACScan flow cytometer (Becton Dickenson, Mountain View, CA, USA) and CellQuest Pro (IVD) software (Becton Dickenson, Mountain View, CA, USA). The assays were done in duplicate.
DAPI staining
To assess the nuclear morphology, KTH-13-t-Bu-treated C6 glioma cells in a 12 well-plate were stained with bisbenzimide solution (Hoechst 33258; Sigma) as reported previously [1516]. Bisbenzimide (0.1 mg/ml) was dissolved in PBS/glycerol (1:1) solution. After rinsing with PBS, the cells were fixed by 4% paraformaldehyde in 0.1 M phosphate buffer and two drops of bisbenzimide solution were added. The cells were observed under a fluorescence microscope with an excitation wavelength of 365 nm.
Preparation of cell lysates and immunoblotting analysis
KTH-13-t-Bu-treated C6 glioma cells (5×105 cells/ml) were washed three times in cold PBS and then lysed in lysis buffer (20 mM Tris-HCl, pH 7.4, 2 mM EDTA, 2 mM ethyleneglycotetraacetic acid, 50 mM β-glycerophosphate, 0.1 mM sodium vanadate, 1 mM dithiothreitol, 2% Triton X-100, 10% glycerol, 2 µg/ml aprotinin, 1 µg/ml pepstatin, 2 µg/ml leupeptin, 1 mM benzimide, 1.6 mM pervanadate, 20 mM NaF, and 50 µM PMSF) for 2 h on ice. The lysates were clarified by centrifugation at 12,000 × rpm for 10 min at 4℃ and then stored at -20℃ until used.
Whole cells were analyzed by immunoblotting [17]. Proteins were separated on 10% SDS-polyacrylamide gels and transferred by electroblotting onto polyvinylidenedifluoride (PVDF) membrane. Membranes were blocked for 60 min in Tris-buffered saline containing 3% FBS, 20 mM NaF, 2 mM EDTA, and 0.2% Tween-20 at room temperature. The membranes were incubated for 60 min with specific primary antibodies at 4℃, washed three times with the same buffer, and then incubated for an additional 60 min with HRP-conjugated secondary antibodies. The total and active caspases (3, 8, and 9) and total and phosphorylated Bax, Bcl-2, STAT3, PI3K/p85, Src, and β-actin were visualized with an ECL system (Amersham, Little Chalfont, Buckinghamshire, UK).
RESULTS
Effect of KTH-13-t-Bu on the proliferation of cancer cells
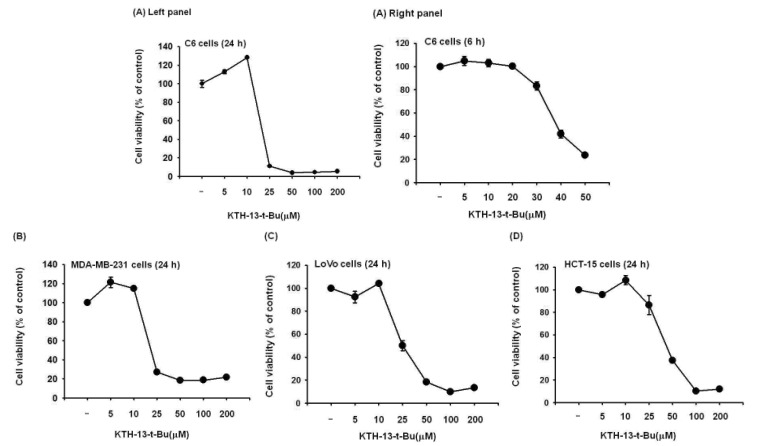
To confirm the effect of KTH-13-t-Bu as an anti-cancer compound, we tested its ability to suppress the proliferation of C6 glioma, MDA-MB-231, LoVo, and HCT-15 cells. As we expected, C6 glioma cell proliferation was dose-dependently decreased by KTH-13-t-Bu (Fig. 2A). Proliferation of other cells was also decreased by KTH-13-t-Bu in a dose-dependent manner (Fig. 2B, 2C, and 2D). Inhibitory levels (IC50 values) of this compound against the proliferation of the cancer cells were summarized in Table 1.
Effect of KTH-13-t-Bu on inducing apoptosis in C6 glioma cells
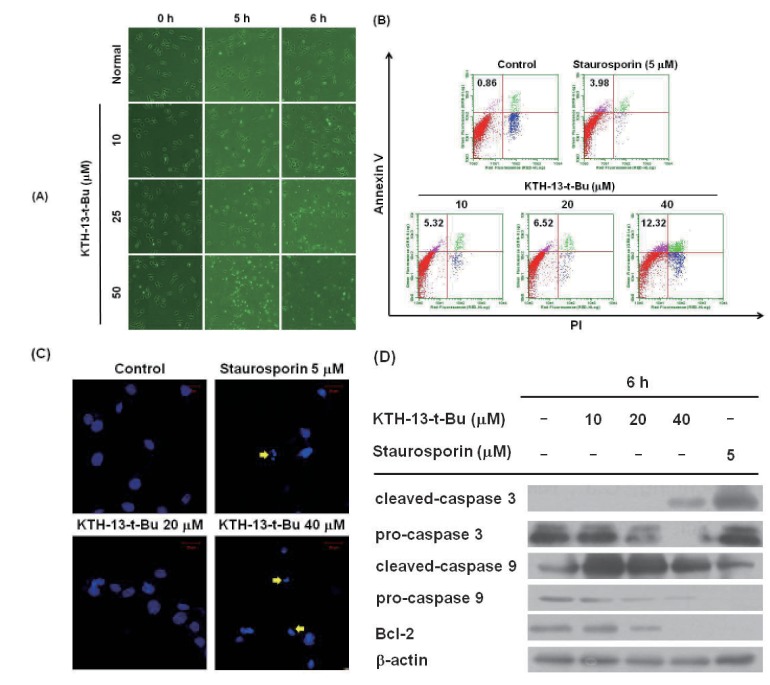
We next examined whether the anti-proliferative activity of KTH-13-t-Bu is derived from its apoptosis-inducing effect in C6 glioma cells. Since strong inhibitory activity of tested cells was displayed in the cells (Table 1) and we used the cell line in our previous experiments with KTH-13 derivatives [18], C6 glioma cells were selected. As shown in Fig. 3A, actin cytoskeleton-dependent morphological changes occurred after incubation with 25 µM KTH-13-t-Bu for 6 h, while 50 µM KTH-13-t-Bu induced morphological changes in 5 h. Annexin V-FITC staining, which indicates early apoptosis, dose-dependently increased from 0.86 to 12.32% (Fig. 3B). Additionally, annexin V-FITC staining increased over time from 0 to 3 h at 40 µM KTH-13-t-Bu (data not shown). As Fig. 3C shows, blebbing, cell shrinkage, and nuclear fragmentation appeared after treatment with KTH-13-t-Bu (40 µM). To identify dose-dependent effects of KTH-13-t-Bu on expression of apoptosis-related proteins, we analyzed the levels of the cleaved and pro-forms of caspase-3 and caspase-9, well-known apoptosis-inducing factors [19], as well as others (Bcl-2, which is apoptosis-preventing protein [20], and β-actin). Interestingly, it was revealed that the pro-forms of caspase-3, caspase-9, and Bcl-2 were decreased by this compound in a dose-dependent manner (Fig. 3D). In parallel, the cleaved forms of caspase-3 and -9 were increased by KTH-13-t-Bu (Fig. 3D), strongly supporting its apoptosis-inducing activity.
Effect of KTH-13-t-Bu on the suppression of cell survival signaling pathway
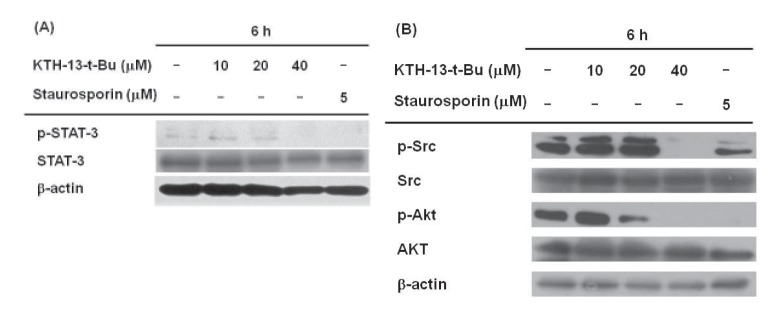
To confirm whether KTH-13-t-Bu affects cell survival signaling, we checked the phosphorylation levels of STAT-3, Src, and AKT, which regulate cell survival. The enzymes were chosen based on our previous finding in which KTH-13 were found to diminish the phosphorylation of these proteins without altering total proteins [18]. As expected, the levels of these phosphoproteins were clearly suppressed by 40 µM of KTH-13-t-Bu. A standard pro-apoptotic compound, staurosporin, also reduced phospho-protein levels of Src, AKT, and STAT-3 (Fig. 4A and 4B), implying that our experimental conditions were well established.
DISCUSSION
Cordyceps species were reported to exhibit anti-cancer activity [2122]. Cordyceps militaris (AECM) induced MDA-MB-231 to undergo apoptosis [23]. Cordyceps pruinosa butanol fraction (Cp-BF) inhibited proliferation and promoted apoptosis in HeLa cells [24]. Previously, we determined that KTH-13 from Cb-BF can also suppress cell growth and viability and induce apoptosis in glioma C6 and MDA-MB-231 cells [7]. We hypothesized that a KTH-13 derivative might have anti-proliferative and pro-apoptotic effects. To confirm this hypothesis, we synthesized KTH-13-t-Bu and tested whether this derivative can reduce the viability of several cancer cell lines including C6 glioma, MDA-MB-231, LoVo, and HCT-15 cells. As we expected, the viability of C6, MDA-MB-231, LoVo, and HCT-15 cells was suppressed by KTH-13-t-Bu in a dose-dependent manner similar to treatment with other natural compounds such as ginsenoside Rg3 and cordycepin [2526]. The IC50 of KTH-13-t-Bu (20 to 40 µM) is presented in Table 1. These results led us to further research its anti-cancer activity.
To investigate the anti-cancer mechanism of KTH-13-t-Bu, we determined the morphological changes in C6 glioma cells treated with KTH-13-t-Bu. Previous studies reported that actin, a cytoskeletal protein, is cleaved by caspases during cell apoptosis [27]. As shown in Fig. 3A, 25 µM KTH-13-t-Bu stimulated morphological changes in C6 glioma cells at 6 h and 50 µM KTH-13-t-Bu at 5 h. These results imply that KTH-13-t-Bu can induce apoptosis in cancer cells. To confirm this effect, we performed annexin V-FITC staining [282930]. Fig. 3B depicts the increase in annexin V-FITC staining level from 0.86 to 12.32% during KTH-13-t-Bu treatment, indicating that KTH-13-t-Bu can promote apoptosis in cancer cells. To confirm the apoptosis-inducing activity of KTH-13-t-Bu, we also assessed nuclear fragmentation by confocal microscopy with DAPI as reported previously [153132]. Fig. 3C shows that KTH-13-t-Bu induced nuclear fragmentation in C6 glioma cells.
The molecular mechanism of the pro-apoptotic effect of KTH-13-t-Bu was examined. Since active caspases have been identified as key enzymes regulating apoptosis [33], the levels of cleaved caspase proteins were detected after treatment with KTH-13-t-Bu. We chose caspase-3 and -9, since they represent mitochondrialdependent intrinsic apoptosis [34]. As shown in Fig. 3D, procaspase-3 and -9 levels decreased dose-dependently while cleaved caspase-3 and -9 increased. Bcl-2, an apoptosis inhibitor [35], also decreased in KTH-13-t-Bu-treated cells. These results indicate that KTH-13-t-Bu induces apoptosis through mitochondrialdependent intrinsic pathways.
Since apoptosis is closely linked to the down-regulation of cell survival signaling, we examined the effects of KTH-13-t-Bu on cell survival signaling. Src, PI3K, and AKT have been reported to have anti-apoptotic effects via various downstream targets, including STAT-3 [363738]. Fig. 4A shows that STAT-3 phosphorylation decreased dose-dependently in response to KTH-13-t-Bu. KTH-13-t-Bu also decreased phosphorylation of Src and AKT (Fig. 4B), which are upstream enzymes for STAT-3 activation [39]. Src phosphorylation affects Bcl-2 activity and cell apoptosis by regulating STAT-3 [40]. These results imply that KTH-13-t-Bu blocks cell survival signaling by inhibiting the Src/AKT-STAT3 cascade.
Our results strongly show that structural change of KTH-13 can give its derivatives more active in view of their anti-cancer activities. Thus, interestingly, simple change in 4-isopropyl group (IC50=64.9 µM) of ring B [18] to tert-butyl group (IC50=21.1 µM) led the compound to the enhancement of cytotoxicity up to 3-fold in C6 glioma cells (Fig. 1 and Table 1), implying that the hydrophobicity of side chain of C4 in ring B might critically contribute to the interaction of KTH-13 analogs with target protein(s). Furthermore, removal of ring C [4-isopropyl-2-(1-phenylethyl) aniline] in KTH-13 backbone has negatively affected to its anti-cancer activity [41], implying that the polarity of ring C seems to be also important to maintain its anti-cancer activity. Therefore, to further improve anti-proliferative activity of KTH-13 derivatives, additional modification of C4 side chain in ring B will be continued by introducing more hydrophobic functional groups. In addition, identification of exact target molecule toward this compound should be also continued to understand its full interpretation of drug's action. Therefore, relevant works will be preceded in the following studies.
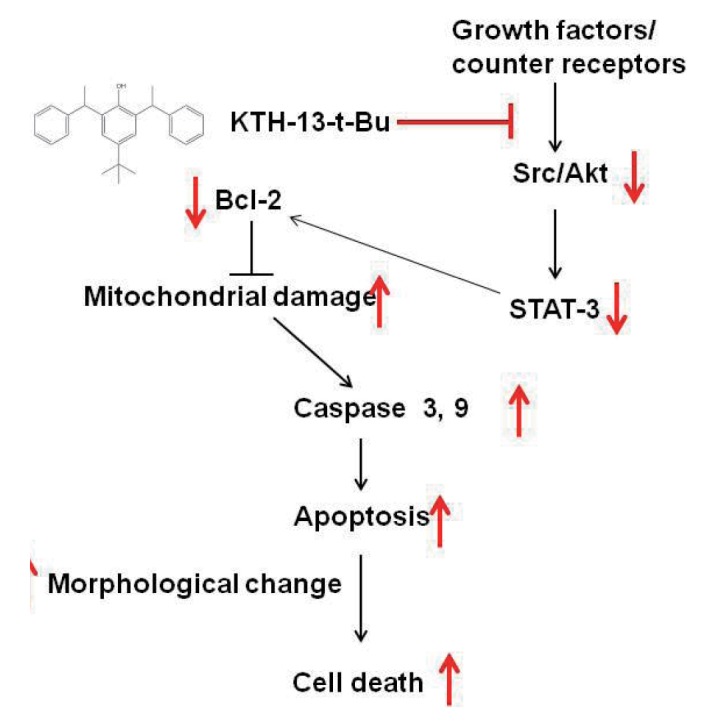
In summary, we demonstrated that KTH-13-t-Bu can suppress cancer cell viability by inducing apoptosis, as summarized in Fig. 5. KTH-13-t-Bu induces pro-apoptotic signaling mediated by caspase and Bcl proteins. Furthermore, KTH-13-t-Bu suppressed Src/AKT-STAT3 signaling, a cell survival cascade. Because KTH-13-t-Bu shows a clear anti-cancer effect, it has the potential to be used for cancer treatment. This study implies that derivatizing compounds from C. bassiana has the potential to increase their anti-cancer activity.

 XML Download
XML Download