

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
CD4+CD25+ regulatory T cells (CD4+CD25+ Tregs) characterized by the expression of the IL-2 receptor α-chain (CD25) show a potent immunosuppressive function and contribute to immunologic self-tolerance by suppressing potentially auto-reactive T cells [1]. Deficiency or dysfunction of CD4+CD25+ Tregs may predispose to autoimmune diseases [2]. In recent years, attention has focused on the forkhead box family transcription factor Foxp3, which is critical for both the identification and function of CD4+CD25+ Tregs [3,4].
Some studies support the observation that CD4+CD25+ Tregs play a role in the control of systemic autoimmune disease, and that the loss of this particular cell population enhances the clinical symptoms of chronic arthritis. Morgan et al. used the collagen-induced arthritis (CIA) murine model to demonstrate that the depletion of CD4+CD25+ T cells before CII immunization greatly hastened the onset and severity of arthritis and markedly increased CII-specific antibodies [5]. Adoptively transferring CD4+CD25+T cells could be used therapeutically in CIA despite a lack of reduction in systemic CII-specific T and B cell responses [6]. CD4+CD25+ Tregs and their role in antigen-induced arthritis (AA) have also been investigated [7]. Depletion of CD25+ cells prior to arthritis induction led to an exacerbation of disease with increased cellular and humoral immune responses. Transfer of CD4+CD25+ T cells into immunized mice at the time of induction of AA decreased the severity of disease, but was not able to cure established arthritis. All of these studies showed that the transferred CD4+CD25+ T cells could be traced to the synovial tissue in affected joints, indicating that these cells may modulate inflammation locally.
Although the frequency of CD4+CD25+ Tregs in joint fluid from rheumatoid arthritis (RA) patients was markedly increased in most previous studies [8,9,10,11,12,13], the CD4+CD25+ Tregs counts in the peripheral blood (PB) of RA patients compared to normal controls remain controversial: some studies report normal numbers [9,10]; some report an increase [11]; and others report a decrease [12,13]. Some of this variability may be explained by differences in the labeling and definition of naturally-occurring CD4+CD25+ Tregs. Some studies used CD4+CD25high T cells, whereas others used the CD4+CD25bright T cells. Besides the differences in detection technology, it may be that the PB obtained by these study groups was most likely taken at different clinical stages of RA, providing an explanation for the conflicting results. Thus, although the importance of CD4+CD25+ Tregs in RA has been emphasized, the dynamic ratios of CD4+CD25+ Tregs at different clinical stages have not been fully characterized. In addition, several reports describing the frequency of CD4+CD25+ Tregs in the PB of RA patients did not take Foxp3 into account [8,10,11,12]. In the present study, we chose the model rat collagen-induced arthritis (CIA), and compared the frequency of CD4+CD25+ Tregs in PB of CIA rats during the development of arthritis with healthy controls, in order to determine the correlation between their frequency and disease progression.
METHODS
Reagents
Type II collagen (CII) was extracted from chicken sternums and purified by our group, as described before [14]. Fluorescein isothiocyanate (FITC) anti-rat CD4, phycoerythrin (PE) anti-rat CD25, FITC anti-rat IgG2b and PE anti-rat IgG1 antibodies were purchased from Caltag Laboratories and the Regulatory T cell Staining Kit #3 was from eBiosciences.
Animals
Male Wistar rats (110~130 g) were obtained from the Experimental Animal Center of Anhui Medical University. They were housed in standard cages at a constant temperature of 22±1℃, relative humidity 55±5% with a natural dark-light cycle for 1 week before the experiment. The animals had free access to food and tap water. The experimental animal protocol was approved by the Anhui Medical University Animal Care and Use Committee.
Establishment of collagen-induced arthritis (CIA)
Collagen-induced arthritis (CIA) was established in Wistar rats as described in our previous paper [15]. Briefly, 2 mg/ml CII was emulsified with an equal volume of IFA. On day 0, rats were injected intradermally with 0.5 ml of the emulsion (containing 0.5 mg of CII) at the base of the tail and other 3~5 sites on the back. Seven days later, a second injection of the emulsion was administered in the same way.
Arthritis assessment
Rats were inspected daily for signs of arthritis characterized by edema and/or erythema in the paws. Bi-hind paw volumes were determined twice a week beginning on the day when arthritic signs were first visible with a volume meter (Shandong Medical Scientific Equipment Station, Shandong, China). The paw swelling (ml) was expressed as an increase in mean bi-hind paw volume by subtracting that at day 0.
Flow cytometric analysis
At a series of time points post-immunization, peripheral blood was collected from angulus oculi medialisin into heparin-containing tubes. The frequency of CD4+CD25+ Tregs was detected by flow cytometry. Briefly, 100 µl whole blood was lysed with NH4Cl lysing solution, mixed thoroughly, and incubated for 8 min in the dark at 4℃. Samples were centrifuged at 500 g for 10 min. The supernatant was removed and washed twice with cold PBS. Then cells were stained with FITC-conjugated anti-CD4 antibody and PE-conjugated anti-CD25 antibody or appropriate isotype control antibody in the dark for 30 min at 4℃. All antibodies were used at concentrations titrated for optimal staining according to the manufacturer's protocol. Samples were centrifuged at 500 g for 10 min. The supernatant was removed and washed twice with cold PBS. Then, cells were incubated with freshly prepared Fixation/Permeabilization working solution overnight at 4℃. After blocking with Fc block in Permeabilization Buffer, cells were then stained with PE-Cy5-conjugated anti-Foxp3 antibody in the presence of Permeabilization Buffer for 45 min in the dark. The labeled cells were detected immediately on a FACS Calibur (Becton-Dickinson, CA, USA) after washing twice with the Permeabilization Buffer. Analysis of 10,000 lymphocyte events per tube was performed using WinMDI software (Joseph Trotter, Scripps Research Institute, La Jolla, CA).
Statistical analysis
Data were expressed as (mean±S.E.M) in figures. Significant differences between groups were determined by Student's t-test. For linear regression analysis, Spearman correlation coefficient was calculated. p-values less than 0.05 were considered significant for all statistical tests.
RESULTS
Signs of arthritis in CIA rats
The onset of arthritis appeared on about day 10 after the injection of CII (three days after the second immunization), with a peak onset on about day 17. During the development of arthritis, CIA rats demonstrated significantly more paw swelling than the normal ones (Fig. 1A). Fig. 1B is the photo of a paw of a normal rat on day 17, and Fig. 1C is the photo of a paw of a CIA rat on day 17.
Proportion of CD4+CD25+ Tregs during the arthritis progress
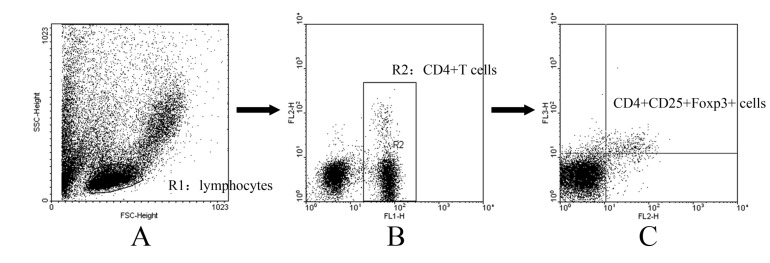
The anti-Foxp3 antibody was used to identify peripheral blood Foxp3-expression lymphocytes in the three-color flow cytometric analysis that also analyzed the CD4 and CD25 expression to correlate the Foxp3 expression, since CD4+ CD25+ Treg cells presumably co-express CD4, CD25 and Foxp3. Gating strategy used to determine the proportion of CD4+CD25+Foxp3+ Tregs was summerized in Fig. 2. Briefly, lymphocytes were gated using an FSC vs. SSC plot (Fig. 2A), followed by a CD4+ gate in a CD4 (FL-1) vs. CD25 (FL-2) plot (Fig. 2B). Then, in CD4+ T cells, Tregs were discriminated according to their CD25 (FL-2) and Foxp3 (FL-3) expression, by which CD4+CD25+Foxp3+ cells are considered as Tregs (Fig. 2C). In our study, Foxp3-expressing cells were barely detected in the CD4-negative lymphocytes, indicating that Foxp3 is predominantly expressed in Fig. 1. the CD4+ T cells in the PB. Therefore, CD4+CD25+Foxp3+ T cells could well represent CD4+CD25+ Tregs.
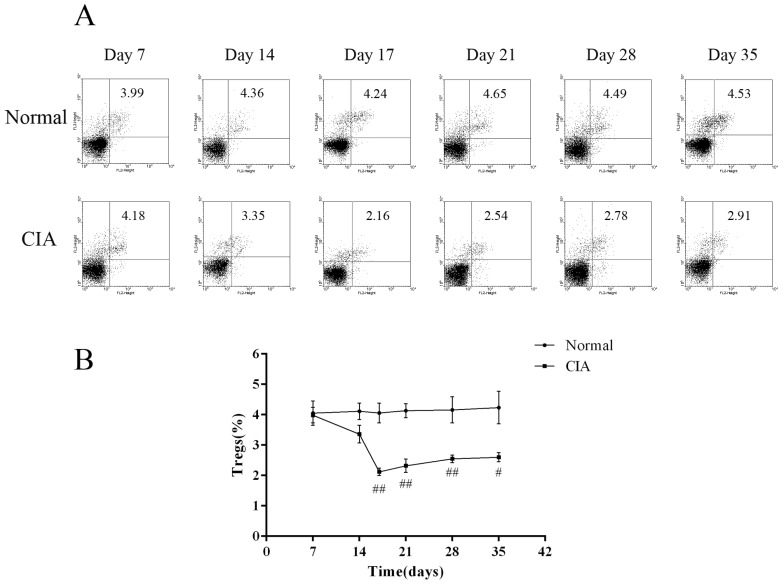
To investigate changes in the number of CD4+CD25+ Tregs as the arthritis progressed, dynamic analysis of the profiles of Foxp3-expressing cells was carried out at a series of time points. The proportion of CD4+CD25+Foxp3+ T cells in normal rats was almost the same during the study period, while it was changing in CIA rats during the progression of arthritis. During the first 2 weeks post-immunization, the percentage of CD4+CD25+Foxp3+ T cells in PB of CIA rats was only a little lower than normal rats, with the difference between the two groups not being significant. However, when the inflammation develops, the ratio was significantly decreased in the immunized group compared to normal rats. On day 17, when the arthritis was most serious, as calculated by paw swelling, the frequency of CD4+CD25+Foxp3+ T cells in CIA rats was the lowest. After that, the ratio rebounded, but is still lower than the normal level (Fig. 3). Therefore, CIA rats showed a significant decrease in CD4+CD25+ Treg percentage during arthritis development when compared to healthy rats.
Correlation between CD4+CD25+ Tregs frequency and the arthritis activity
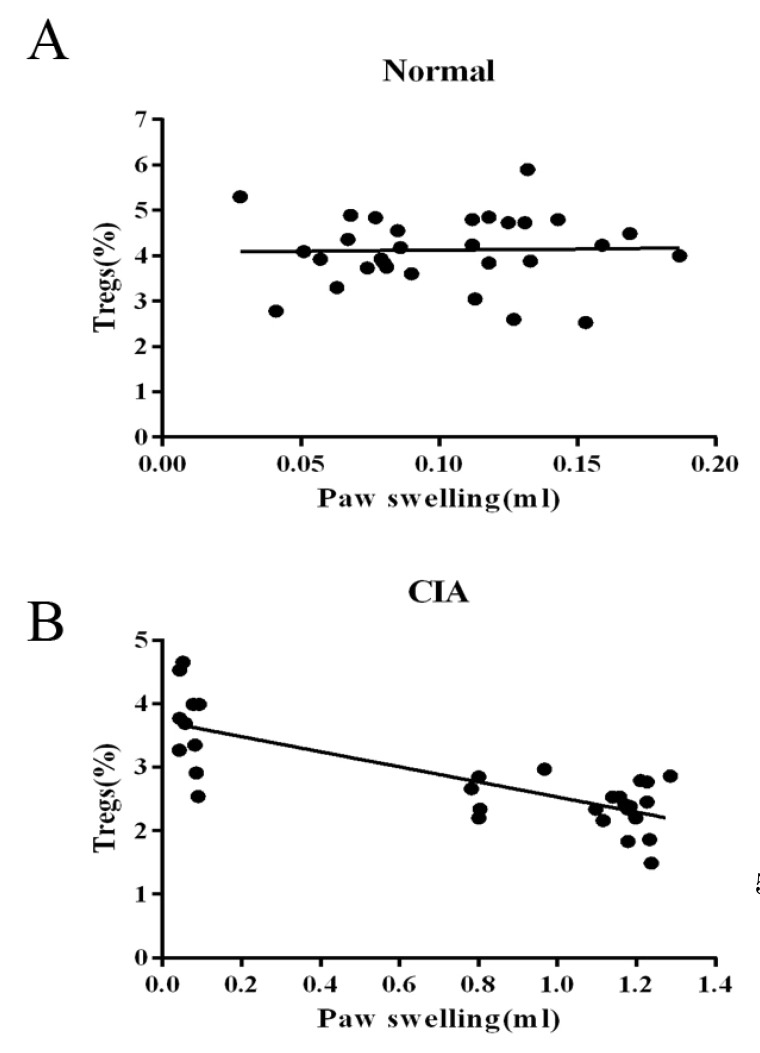
We then determined by simple linear regression whether the frequency of PB CD4+CD25+ Tregs is involved in the immunoregulation of this disease. As the sample size of flow cytometric analysis was smaller than that used for the paw volume measurement, we only chose data from the rats that had been determined by both of the two targets. As shown in Fig. 4, the relationship between the frequency of CD4+CD25+Foxp3+ T cells populations in CD4+ T cells and immune activation was not found in normal rats, while the frequency was negatively correlated with paw swelling with statistical significance (r=-0.786) in CIA rats.
DISCUSSION
CIA is an autoimmune-mediated polyarthritis that shares important similarities with RA in histology and immunology [16,17]. The immune reaction of CIA rats is regulated by complex mechanisms, which are incompletely understood, but regulatory T cells are able to suppress the activation of CD4+ T cells [18]. CD4+CD25+ Tregs are a subset of T cells which are involved in peripheral immune tolerance by suppressing auto-reactive T cells. Their role in autoimmune disease, which occurs through a breakdown of tolerance, is of particular interest in trying to ascertain the mechanisms of disease progression [19]. Several reports have investigated the expression of CD4+CD25+ Tregs, but the relationship between the frequency of CD4+CD25+ Tregs and disease progression has not been fully described, especially among CIA rats. In the present study, we studied the correlation of the alternation of CD4+CD25+ Tregs in the PB of CIA rats and disease progression.
The identification of CD4+CD25+ Tregs during ongoing immune responses is complicated, as activation markers such as CD25 are also expressed on activated effector T cells [20,21]. Over the past few years, several studies have demonstrated that the intracellular marker Foxp3 is expressed in CD4+CD25+ Tregs and therefore it appears to be of great significance for both the identification and function of CD4+CD25+ Tregs [22]. Additionally, Foxp3 expression is limited to CD4+CD25+ Tregs in mice and could not be induced in CD4+CD25- effector cells upon activation [3]. Thus, we characterized the CD4+CD25+ Tregs as the percentage of CD4+CD25+Foxp3+ T cells among CD4+ T cells. Our results showed that the frequency of CD4+CD25+Foxp3+ T cells was the same as that in the controls 2 week post-CII-immunization. At the chronic stage of inflammation, the percentage in CIA rats significantly decreased, with the expression of Foxp3 showing a gradual decrease along with arthritis progression. As shown in Fig. 3, the frequency of CD4+ CD25+Foxp3+ T cells in CIA rats was significantly lower than that of normal rats after day 17. We also observed that the down-regulation of the frequency of CD4+ CD25+ Tregs was significantly correlated with paw swelling in CIA rats (r=-0.786, p<0.01), suggesting that the continuous arthritis may be due to the decrease of CD4+CD25+ Tregs over the course of disease progression. In contrast, no relationship between the frequency of CD4+CD25+ Tregs and immune activation was seen in normal rats (p>0.05). We also found that the mRNA and protein level of Foxp3 was decreased in the spleens of CIA rats than in normal controls and could be improved by leflunomide both in vivo and in vitro [23]. These results are consistent with our hypothesis of the involvement of CD4+CD25+Tregs in CIA progression and with the findings of several published studies, which showed an increase of CD4+CD25+Tregs in the therapeutic effect of anti-arthritic drugs on CIA [24,25,26,27].
Taken together, our data indicate that CD4+CD25+ Tregs might be involved in the immune modulation during the course of CIA, which enhances the current knowledge of mechanisms of the immune disorder in CIA. It should be noted, however, that much of the data regarding CD4+CD25+ Tregs has been generated using animal models. Thus, future studies will examine the dynamic frequency of CD4+CD25+ Tregs at different clinical stages to provide new insights into the autoimmune genesis and therapeutic strategies of RA. It is hoped that by understanding the role of CD4+CD25+ Tregs in autoimmunity, a reliable therapy may be developed to cure the disease.

 XML Download
XML Download