

 PDF
PDF ePub
ePub Citation
Citation Print
Print


ABBREVIATIONS
MMC
migrating motor complex
ICC
Interstitial cell of Cajal
ICC-MY
ICC in the myenteric region
ICC-SEP
ICC in line septa
CM
circular muscle
LM
longitudinal muscle
KRB
Krebs-Ringer solution
RMP
resting membrane potential
AUC
area under the curve
L-NA
Nw-oxide-L-arginine
NO
nitric oxide
SNP
sodium nitroprusside
INTRODUCTION
Small bowel motility consists of spontaneous and rhythmic phasic contractions from slow waves and periodic mass movements as migrating motor complexes (MMC) in animal models. Phasic contractions mix materials in the intestine, and MMC propel the materials from the proximal to distal intestine. Slow waves originate from the interstitial cells of Cajal (ICC), especially from the ICC in the myenteric region (ICC-MY) between circular muscle (CM) and longitudinal muscle (LM) in the small bowel. The electrical activities of the ICC, which include changes in membrane potential from the activation of various ion channels, provoke slow waves in smooth muscle cells connected with ICC by gap junctions [1234]. Thus, the ICC acts as a pacemaker for the generation of smooth muscle phasic contractions. However, MMC are thought to be controlled by the enteric nervous system and not by the ICC. The MMC have been reported to be subjected to cholinergic agents or neuronal blockers [5678].
Although some reports observed slow waves and contractions in the smooth muscle of the human small intestine [910111213], the mechanisms of electric or contractile activities remain unknown. More complex mechanism should be involved in conduction of the slow wave from the ICC to the smooth muscle in human, which might be different from the small animal. A recent study using calcium imaging techniques reported that the ICC in the human jejunum could generate spontaneous pacemaker activity that actively propagates to smooth muscles [14], and ICC-MY as well as ICC in line septa (ICC-SEP) in the CM layer are thought to have important roles in effective conduction through thick human smooth muscle [15]. We assumed that if the slow waves can be conducted sufficiently from ICC-MY to the CM close to the myenteric region and eventually to the CM near the submucosa, both myenteric and submucoal side of CM has no differences in electrical activities of membrane potential.
We performed this study to explore the motility of the human ileum, evaluating the electrophysiological characteristics of smooth muscle, slow waves conducted from the ICC to smooth muscles and the mechanical characteristics of smooth muscle strips and ileal segments in vitro.
METHODS
Tissue acquisition
Human ileum was obtained immediately from patients undergoing ileal resection for non-obstructive bowel diseases. As the amount of ileal tissue needed for the experiment was very small, there was no further resection of the bowel than for routine operations. This study was approved by the Institutional Review Board of the Clinical Research Institute of the Seoul National University Hospital (IRB approval number: H-0603-071-170).
After resection of the bowel, a 4×2-cm ileal segment was removed from the normal ileal region within 30 cm of the ileocecal valve. Specimens were immediately placed into oxygenated Krebs-Ringer solution (KRB). The KRB contained (in mM) 120.4 NaCl, 5.9 KCl, 15.5 NaHCO3, 11.5 glucose, 1.2 MgCl2, 1.2 NaH2PO4 and 2.5 CaCl2. This solution had a pH of 7.3~7.4 at 37.5℃ when bubbled to equilibrium with 97% O2~3% CO2.
Tissue preparations
1. Preparation for electrophysiological intracellular recording of membrane potential in cross-sectional CM
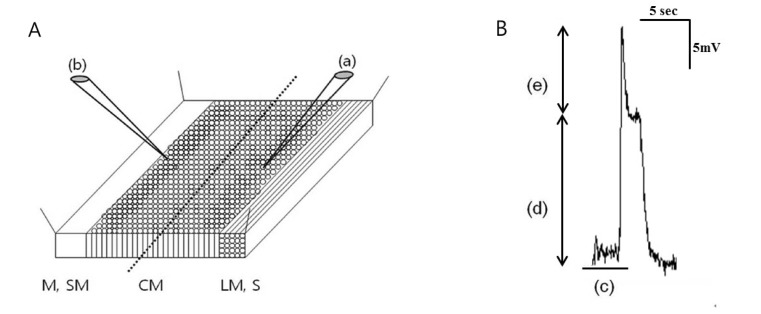
Tissues were transferred to a Petri dish coated with Sylgard (Dow corning Co., USA) and pinned downed in a dissecting dish. The muscles were cut parallel to the LM fibers with a knife consisting of a pair of sharp parallel scalpel blades set 1.5 mm apart and turned on their sides to expose a cross section of the entire muscle layers in the electrophysiological chamber. The chamber was constantly perfused with pre-warmed, pre-oxygenated KRB solution. The temperature was maintained at 36.5±0.5℃. The muscles were equilibrated for at least 1 hour before beginning the experiments. Conventional microelectrode recordings were performed using sharp microelectrodes filled with 3 M KCl. Because the tissues were pinned down in a cross-sectional manner, we could differentiate the CM as outer CM (near the myenteric region) and inner (near the submucosal side) and (Fig. 1A). The membrane potential of the outer and inner CM was measured with a high-input resistance electrometer, and outputs were displayed on an oscilloscope. Resting membrane potentials were measured using the pClamp softwareR (version 9.0. Axon Instruments, Foster City, CA, USA) and Origin Software (MicroCal Software, Northampton, MA, USA) program. Resting membrane potential (RMP) (mV), plateau amplitude (mV), upstroke amplitude (mV) and frequency (/min) of the slow waves were analyzed (Fig. 1B).
2. Preparation for mechanical tension recordings of muscle strips
Tissues were pinned down on a Petri dish coated with Sylgard with the mucosa side facing upward. The mucosal and submucosal layers were gently removed with a pair of scissors. CM and LM bundles were obtained by sharp dissection. The size of the muscle strips was 2 mm in width and 1 cm in length.
For recording contractile activities, the muscle strips were attached to an isometric strain gauge (World precision Instruments, Sarasota, FL, USA) with a suture in a tissue chamber perfused with pre-warmed, pre-oxygenated KRB solution. The temperature was maintained at 36.5±0.5℃. The muscle strips were equilibrated for at least 1 hour before beginning the experiments, and a resting force of 9.8 mN (1 g) was applied. The mechanical signals were digitized and recorded on Acknowledge software (Biopac Systems, Inc., Goleta, CA) for data analysis. The frequency (/min), amplitude (mN), and area under the curve (AUC, sec×mN/wave) of the contractions were analyzed. AUC was defined as the integrated area under a single wave.
3. Preparations for mechanical tension recordings of ileal segments
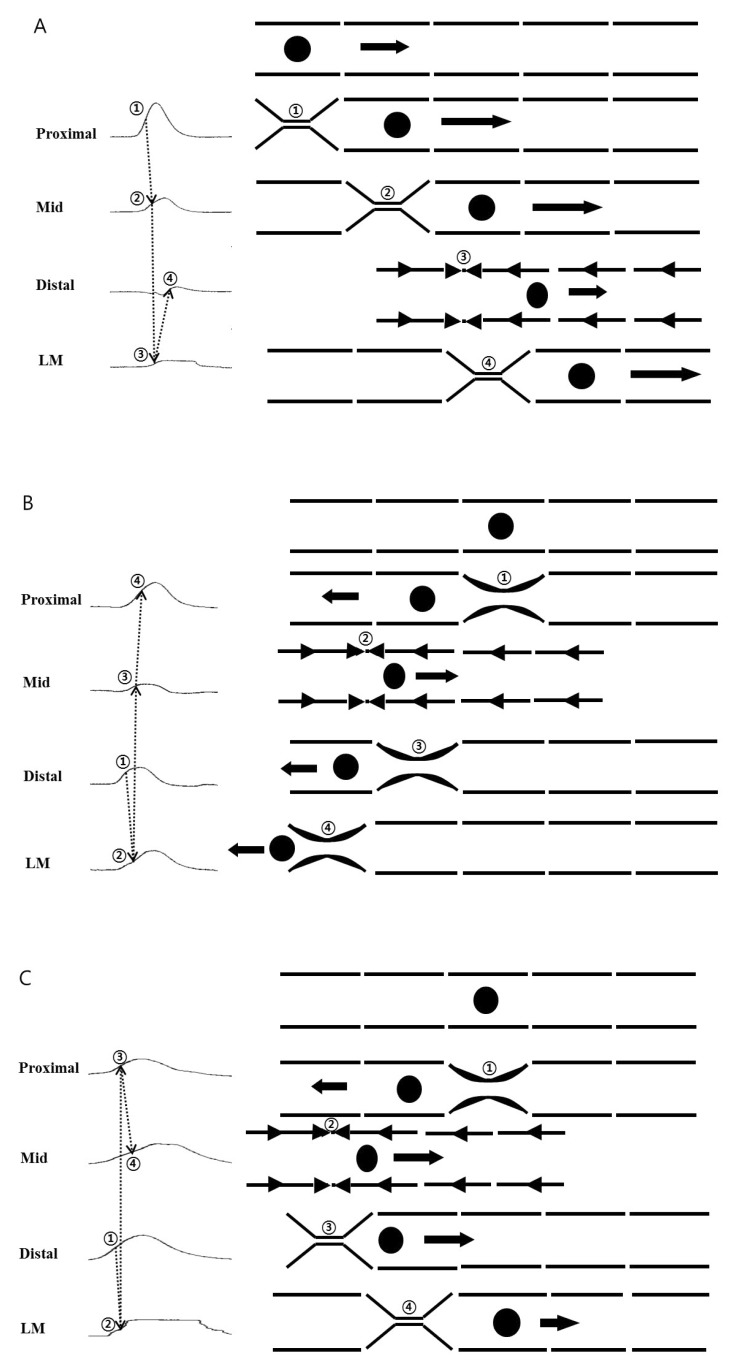
Ileal segments were prepared by cutting the whole layer of the segment parallel to the longitudinal muscle. The size of the segment was 4 cm in length and 2 cm in width. A stainless steel rod was placed parallel to the longitudinal muscle and placed in an organ bath. CM tension was recorded at three (proximal, middle, and distal) sites, and LM tension was recorded by perpendicular traction with sutures placed at each site. Ileal segments were suspended in a tissue chamber perfused with pre-warmed, pre-oxygenated KRB solution. The temperature was maintained at 36.5±0.5℃. The muscles were equilibrated for at least 1 hour before the experiments were begun. A resting force of 9.8 mN (1 g) was applied to each measuring site. The tension of each site was transformed to an isometric strain gauge. The frequency (/min), amplitude (mN), and AUC (sec×mN/wave) of the contractions were measured using Acknowledge software. The propagation patterns were evaluated by analyzing sequences of contractions detected at each site during recording. The sequence was determined by evaluating the half maximal amplitude points on the rising of each contraction. The patterns of propagation were classified as antegrade, which describes propagation from the proximal to distal site, and retrograde, which describes propagation from the distal to proximal site. A mixed pattern of propagation was also defined as contractions without consistent sequences. For evaluating the propagations in a recording of the ileal segment, we analyzed 10 propagations of waves.
Drug treatments
After regular waves were detected in each type of experiment, drugs affecting the enteric nervous system were perfused into a tissue chamber, and changes in the variables were measured. Atropine (1 µM) was used as an anticholinergic drug. Nw-oxide-L-arginine (L-NA) (100 µM), a nitric oxide (NO) synthase inhibitor, and sodium nitroprusside (SNP) (100 µM), an NO donor, were also administered. All drugs were obtained from Sigma Chemical Co. (St Louis, MO, USA).
Immunohistochemistry
Immunohistochemistry was performed to analyze the distribution of ICC in the human ileum using a conventional labeled streptavidin-biotin-peroxidase method (LSAB Kit, DAKO, Glostrup, Denmark) according to the manufacturer's protocol. The primary antibody used was c-Kit (CD117) (DAKO, Glostrup, Denmark, A4502, 1:300). Slides were incubated in biotinylated goat anti-mouse/rabbit immunoglobulin and then in a solution of streptavidin-biotin complex. Immunoreactivity was visualized using 3,3-diaminobenzidine.
RESULTS
Electrophysiological intracellular recordings of membrane potential
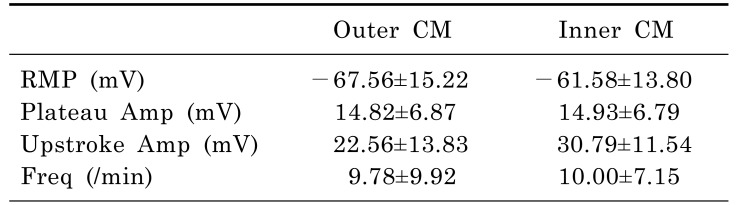
Intracellular recordings of membrane potential were performed in tissues from 10 patients. The mean patient age was 69.20±12.65 years, and the male:female ratio was 6:4. The slow waves were detected immediately after impalement of the outer and inner CM by intracellular recording (Fig. 2A). The RMP of slow waves at outer CM was -67.56±15.22 mV and plateau and upstroke amplitude were 14.82±6.87 and 22.56±13.83 mV, respectively. These did not differ significantly at inner CM. The frequency of slow waves was also not different between outer and inner CM (9.78±9.92 vs 10.00±7.15/min, p=0.959). Detailed analyses of the electrical activities of the outer and inner CM are presented in Table 1.
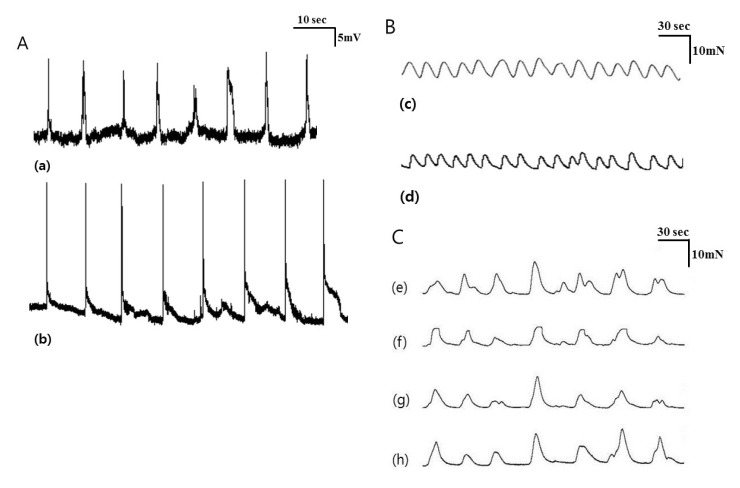
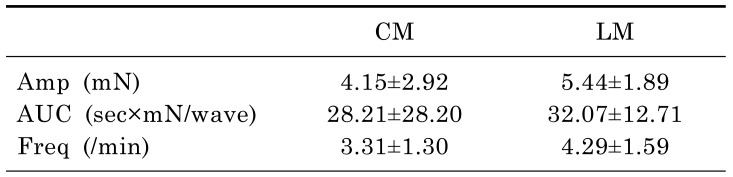
Mechanical tension recordings of smooth muscle strips
Muscle strips obtained from 10 patients were used for tension recordings. The mean age of the patients was 63.00±14.02 years, and the male:female ratio was 5:5. The waves of contraction were detected by tension recordings in the CM and LM (Fig. 2B). The amplitude and AUC at the CM were 4.15±2.92 mN and 28.21±28.20 sec×mN/wave. These did not differ significantly at the LM. The frequency of contractions between the CM and LM were also not statistically significant (3.31±1.30 vs 4.29±1.59/min, p=0.074). Detailed analyses of the data are described in Table 2.
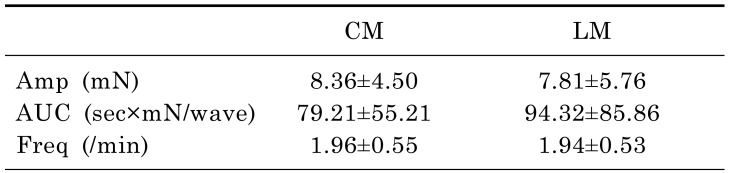
Mechanical tension recordings of ileal segments
Tension recordings were also performed in ileal segments from 12 patients. The mean age of the patients was 60.50±12.41 years, and the male:female ratio was 6:6. The waves of contraction were detected by tension recordings in the proximal, middle, and distal CM and LM. The contractions of the CM nearly always occurred in conjunction with contractions of the LM at a ratio of 1:1 (Fig. 2C). The amplitude and AUC at the CM were 8.36±4.50 mN and 79.21±55.21 sec×mN/wave. These did not differ significantly at the LM. The frequency of contractions were also not statistically significant between the CM and LM (1.96±0.55 vs 1.94±0.53/min, p=0.722). Detailed analyses of the data are described in Table 3.
There were three propagation patterns of contractions in the ileal segment. Antegrade propagation was observed 4.00±1.95 times, and retrograde propagation was observed 2.58±1.73 times per 10 propagations of waves. Mixed patterns were identified 3.42±2.23 times (Fig. 3).
The effects of drugs
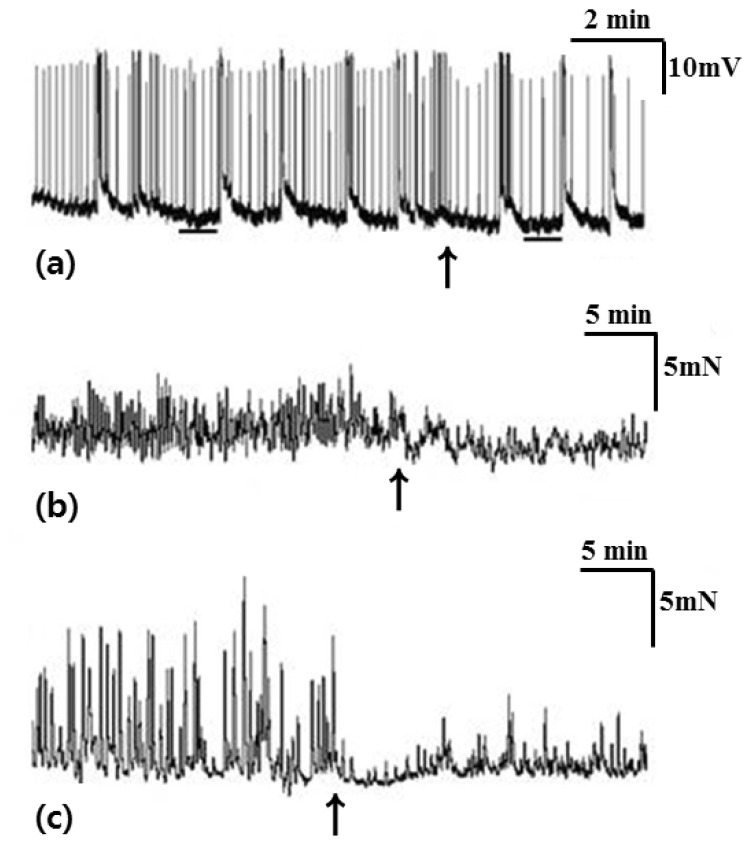
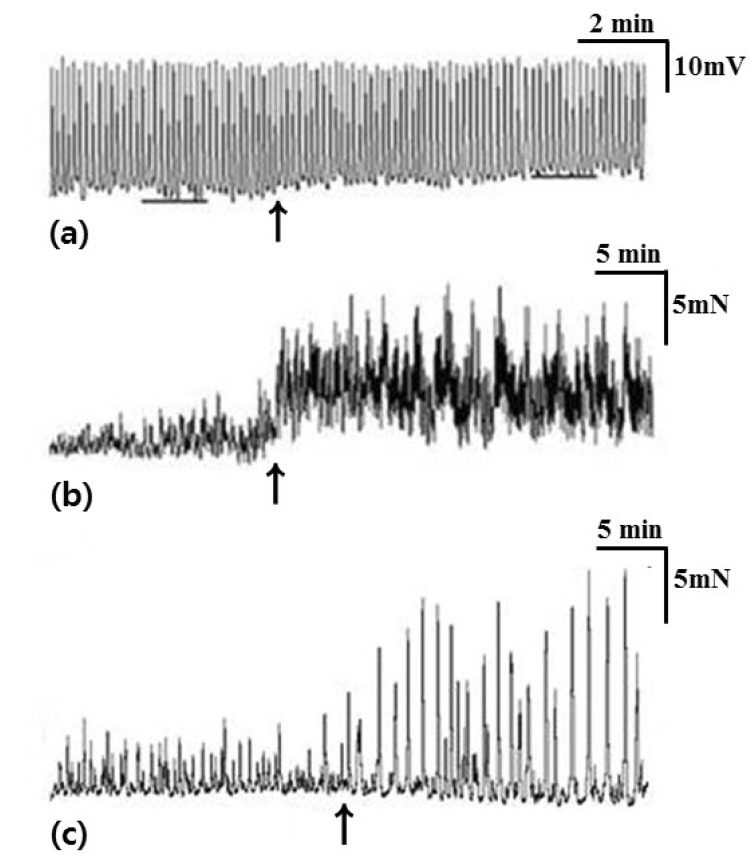
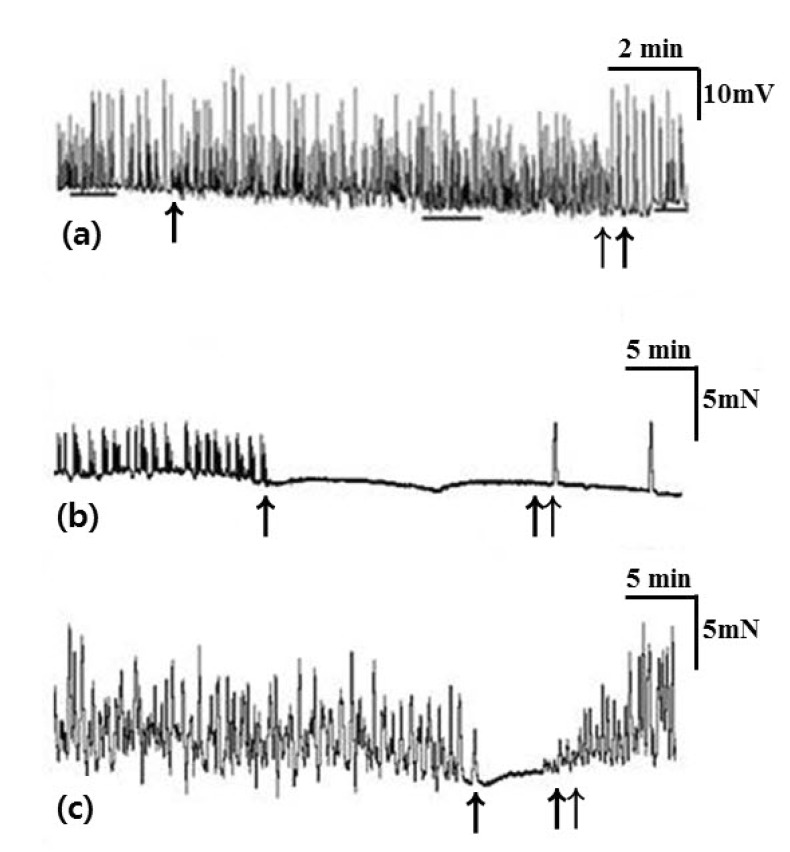
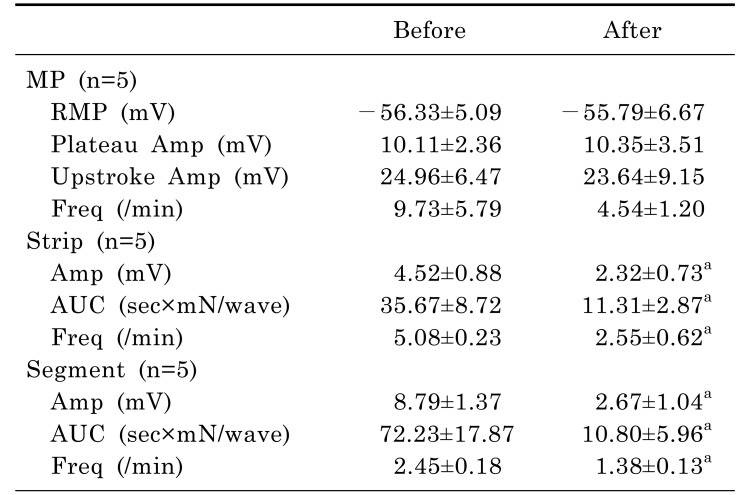
In tension recordings of muscle strips and ileal segments, we observed significant attenuation of the contractile activities after treatment with atropine (Table 4, Fig. 4). In smooth muscle strips, the amplitude of waves of contraction was 4.52±0.88 mN before administration of atropine and 2.32±0.73 mN after administration of atropine (p=0.042). The AUC was 35.67±8.72 sec×mN/wave before administration of atropine and 11.31±2.87 sec×mN/wave after administration of atropine (p=0.043). The frequency of contractions was 5.08±0.23/min before administration of atropine and 2.55±0.62/min after administration of atropine (p=0.043). In ileal segments, the amplitude of waves of contraction was 8.79±1.37 mN before administration of atropine and 2.67±1.04 mN after administration of atropine (p=0.043). The AUC was 72.23±17.87 sec×mN/wave before administration of atropine and 10.80±5.96 sec×mN/wave after administration of atropine (p=0.043). The frequency of contractions was 2.45±0.18/min before administration of atropine and 1.38±0.13/min after administration of atropine (p=0.039).
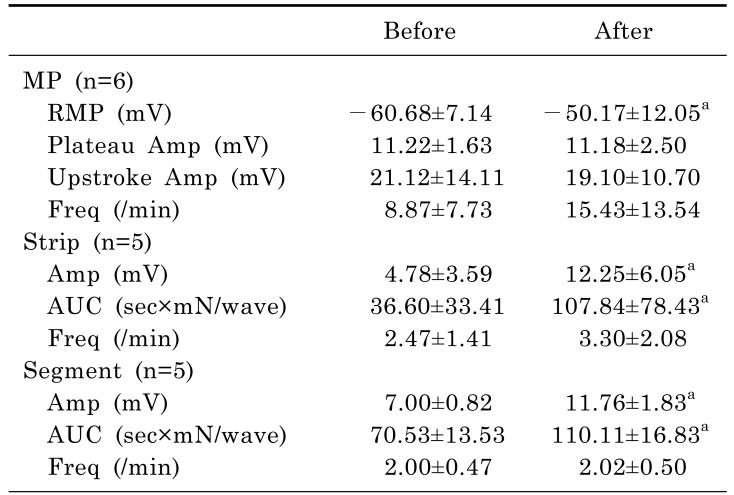
L-NA significantly depolarized the membrane potential of CM and significantly enhanced the contractile activities of the muscle strips and ileal segments (Table 5, Fig. 5). In intracellular recordings, RMP was -60.68±7.14 mV before administration of L-NA and -50.17±12.05 mV after administration of L-NA (p=0.018). In smooth muscle strips, the amplitude of waves of contraction was 4.78±3.59 mN before administration of L-NA and 12.25±6.05 mN after administration of L-NA (p=0.043). The AUC was 36.60±33.41 sec×mN/wave before administration of L-NA and 107.84±78.43 sec×mN/wave after administration of L-NA (p=0.043). In ileal segments, the amplitude of waves of contraction was 7.00±0.82 mN before administration of L-NA and 11.76±1.83 mN after administration of L-NA (p=0.043). The AUC was 70.53±13.53 sec×mN/wave before administration of L-NA and 110.11±16.83 sec×mN/wave after administration of L-NA (p=0.043).
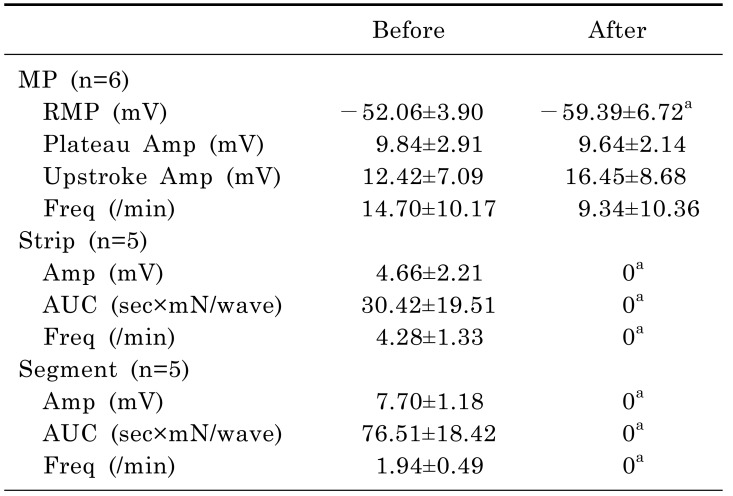
SNP induced significant hyperpolarization of the CM. In tension recordings of muscle strips and ileal segments, significant attenuation of contractile activity was observed after treatment with SNP (Table 6, Fig. 6). In intracellular recordings, RMP was -52.06±3.90 mV before administration of SNP and -59.39±6.72 mV after administration of SNP (p=0.028). In smooth muscle strips, the amplitude of waves of contraction was 4.66±2.21 mN before administration of SNP and 0 mN after administration of SNP (p=0.043). The AUC was 30.42±19.51 sec×mN/wave before administration of SNP and 0 sec×mN/wave after administration of SNP (p=0.043). The frequency of contractions was 4.28±1.33/min before administration of SNP and 0/min after administration of SNP (p=0.043). In ileal segments, the amplitude of waves of contraction was 7.70±1.18 mN before administration of SNP and 0 mN after administration of SNP (p=0.043). The AUC was 76.51±18.42 sec×mN/wave before administration of SNP and 0 sec×mN/wave after administration of SNP (p=0.043). The frequency of contractions was 1.94±0.49/min before administration of SNP and 0/min after administration of SNP (p=0.043).
Immunohistochemistry
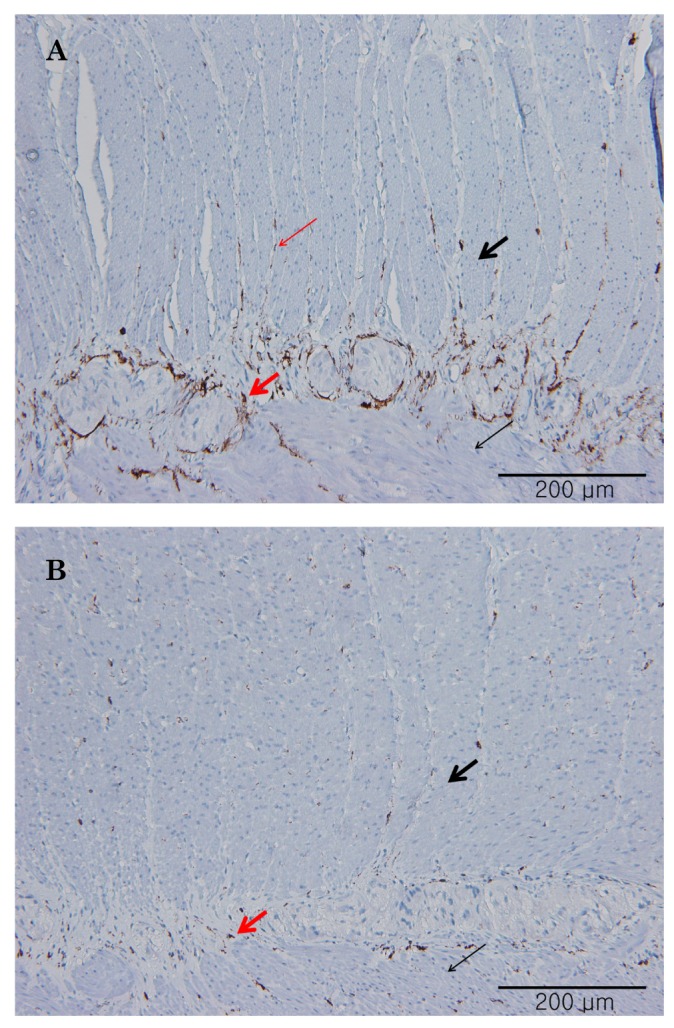
The ICC in the ileum were abundantly distributed in both the myenteric region (ICC-MY) as well as in the septum of CM (ICC-SEP) (n=5). However, ICC-MY and ICC-SEP were not abundant in human colon (n=5) (Fig. 7).
DISCUSSION
There have been few studies to verify human small bowel motility, in detail. We studied the human ileal electrophysiological and mechanical characteristics at three different levels. Electrophysiological activities could be recorded in human ileal smooth muscle cells as slow waves along with RMP, plateau and upstroke amplitude and frequency in the CM in this study. There were no cross-sectional differences in the electrical activities between the outer and inner CM. The slow wave amplitude and frequency did not differ. Upstroke amplitude in inner CM was slight higher than inner CM, but there was no statistical difference. Although we had performed the study with a small sample size and there could be statistical variability, it is possible that the pacemaker potential, generated from ICC-MY, could conduct effectively from the outer CM, close to the ICC-MY, to the inner CM. Immunohistochemical staining supported this theory by demonstrating the anatomical distributions of c-Kit (+) ICC-MY and ICC-SEP.
Electrophysiological and mechanical activities of ileal smooth muscle and ICC
The contraction of small intestinal smooth muscle results from binding of actin and myosin. This is initiated by calcium-mediated phosphorylation of the myosin light chain (MLC20), and activation of myosin light chain kinase (MLCK) by calcium binding calmodulin can phosphorylate the MLC20. Increase of intracellular calcium is the first step of contraction, and it can be due to release of calcium from intracellular stores of sarcoplasmic reticulum and influx of extracellular calcium through voltage-gated calcium channel. Depolarization of membrane potential make the calcium channel open, and it can develop action potentials of slow waves [16].
However, the electrical activity of smooth muscle in the small bowel is attributed to changes in membrane potential secondary to the activation of ion channels and calcium currents in the ICC [117]. Recent studies involving murine small intestine have revealed that pacemaker mechanisms for generation of slow waves are related to the release of calcium from inositol 1,4,5-triphosphate (IP3) receptor-operated stores, the reduction of local calcium due to calcium uptake by mitochondria, unitary potential from non-selective cation channels, and voltage-dependent, dihydropyridine-resistant calcium inward currents [181920]. These pacemaker potentials conduct to smooth muscle cells through electrical coupling by gap junctions, thereby causing the development of slow waves with the same frequency between ICC-MY and smooth muscle cells [21]. Because of voltage dependent L-type calcium channel is involved in the slow wave of smooth muscle cells, which is not in the ICC, upstroke spike amplitude seems to be obvious [22]. Although the smooth muscle in human small intestine cannot generate the slow wave spontaneously, the slow waves can be conducted to the smooth muscle by the gap junction and we could evaluate the function of the ICC indirectly from intracellular recordings of slow waves in the smooth muscle. The two-dimensional network system with electrical coupling between ICC-MY and smooth muscle cells by gap junctions is well established to be critical for the conduction of pacemaker potentials and the generation of slow waves in small mammals [2023]. Although this pacemaker potential could spread rapidly, the waves can propagate only a short distance, approximately 100~200 µm [15]. Therefore, a more complex system is most likely required for the conduction of these potentials in the relatively thick small bowel of larger animals. Previous studies on canine gastric antrum and colon verified the existence of a 3-dimensional network among ICC-MY, smooth muscle cells, and ICC-SEP separating CM bundles [2425]. A recent study using calcium current imaging techniques revealed that the pacemaker potentials from ICC-MY conduct to ICC-SEP and effectively activate the smooth muscle bundles. Because the frequency of the calcium current in the ICC-SEP was observed to be smaller than in ICC-MY, it was suggested that there could be a threshold potential for the activation of ICC-SEP [15]. Additionally, many studies suggested that action potential complexes would exceed the mechanical threshold and the accumulated upstroke spike potentials would result in slow waves for the contraction of smooth muscle [2627]. Our results might support the threshold theory indirectly, as the frequencies of contractions in CM strips or segments were smaller than the membrane potential changes of the outer CM, which were 10.00±2.26, 3.40±0.45, 0.94±0.18/min, respectively. Although the tissues used in intracellular and tension recordings were not exactly matched, those might be considered the differences of frequencies in each level.
The changes of contractile activities by neurotransmitters related to enteric nervous system
The differences of frequency in this study suggested that the contractions could be related to not only slow waves of ICC but also enteric nervous system. MMC have been observed in murine small and large intestine with lower frequency than phasic contraction or slow waves [8]. MMC can occur spontaneously in mutant murine models without ICC-MY and can be controlled by enteric neuronal mechanisms [6]. Furthermore, neurotransmitters from enteric neurons can affect contractions of human gastrointestinal smooth muscles [91112]. Atropine prevents contractions of gastrointestinal smooth muscle by blocking the acetylcholine receptor [8]. Although ICC have been reported to express several receptors for acetylcholine, the generation of pacemaker potential is not be affected by anticholinergic neurotransmitters [91428]. The noncholinergic and nonadrenergic neurotransmitter NO also exhibits inhibitory activity on smooth muscle contractions and affects calcium concentrations in the ICC and smooth muscle cells [293031]. In this study, the RMP was altered, and the contractions of muscle strips and ileal segments were affected by the drugs related neurotransmission. These results indicated that the mechanical characteristics of smooth muscle contractions in human ileum can be controlled by both the electrophysiological mechanisms of slow waves as well as the enteric nervous system. There might be different action mechanisms and relations to provoke activation and contractions at each level of ICC, smooth muscle cells and ileal segment and further study is necessary to elucidate these characteristics of human ileal motility.
The differences of motility between human ileum and colon
Colonic motility also consists of contractions from slow waves and MMC [323334]. However, slow waves are known to be generated from ICC in the submucosa (ICC-SM) and act with myenteric potential oscillation (MPO) from ICC-MY [35]. We have previously reported the electromechanical characteristics of human colon and observed cross-sectional differences between the frequencies of membrane potential changes in the inner and outer CM and LM, unlike our observations in the ileum [36]. ICC-SM, close to the inner CM, might be more important in generating slow waves in the human colon. Additionally, there were significant differences in the contractile activities and the propagation patterns of the right and left colon. The retrograde propagations were more frequently observed in the right colon and might be associated with the mixing function in the right colon, as in the ileum. Comparing the electromechanical characteristics of the ileum with the colon, the frequency of slow waves and the frequency of contractions of smooth muscle were higher. However, the amplitude and AUC of contractions were lower in human ileum (Table 7). These results suggest that the human ileum contracts more frequently with less strength than the colon and might be more suitable to the function of mixing the materials in the bowel.
The ileum has many functions as a gastrointestinal organ, including mixing materials for absorption and moving materials to the colon. Contractions and propagation are critical to perform these functions. We observed cooperative contractions of the CM and LM and equal frequencies of antegrade and retrograde propagation patterns by analyzing the propagation patterns of human ileal segment. Mixed patterns of propagation were similarly observed and may indicate the dynamic motions of the human ileum, involved in both the mixing and propagation functions of the ileum. Our study is unique because analyzing the propagation patterns at human ileal segment might be important as those will be a tool for evaluating the function of small intestine at the bowel motility disorders and a reference for further researches.
Limitations of the study
This study was limited to a small number of experiments in vitro. However, studies on bowel motility using normal human ileum have been rare, and detailed electrophysiological and mechanical characteristics, including the propagation patterns, were particularly investigated in this study. The pathophysiology of diseases exhibiting abnormal bowel motility, such as pseudo-obstruction, small bowel stasis or irritable bowel syndrome, is poorly understood. Recent studies reported that these diseases might be related to abnormalities of MMC or slow wave activities [37]. Further research is necessary to illuminate the pathophysiology of various motility disorders, and our results can be used for reference values of human ileal motility.
Conclusions
In conclusion, there were no cross-sectional differences in the slow waves of the human ileal CM. The differences of frequency of slow waves and contractions in smooth muscle strips and ileal segments indicated that there may be different action mechanisms to provoke activation and contractions at each level. Contractions of the CM were linked with the LM in the tension recordings of the ileal segments, and diverse directions of propagation could represent both the mixing and transport function of the human ileum.

 XML Download
XML Download