

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
In the visual cortex, induction of long-term synaptic plasticity and ocular dominance (OD) plasticity decline with development [1234]. The mechanisms underlying this decline of plasticity have been extensively studied. Increase in γ-aminobutyric acid receptor type A (GABAAR)-mediated inhibition appeared to be an important determinant [56]. Changes in the subunit composition of N-methyl-D-aspartic acid receptor (NMDAR) could also be involved [7]. Another factor which could affect the decline of plasticity may be the changes in extracellular matrix [8]. All these changes are thought to participate in the decrease in synaptic plasticity. This assumption could be supported by the studies demonstrating that manipulations for enhancing plasticity in aged animals are accompanied with the changes in GABAAR-mediated inhibition, NMDAR properties, and extracellular matrix [91011]. These studies on methods to enhance plasticity in aged animals have helped to understand the underlying mechanisms in the developmental decrease in the plasticity and will provide important insights for the treatment of neurodevelopmental diseases [12].
Serotonin [5-hydroxytryptamine (5-HT)] regulates the development of neuronal network [1314] and modulates neurotransmission [15]. It also regulates the induction of long-term synaptic plasticity [1416] and OD plasticity [17]. In juvenile rats, 5-HT suppressed the induction of long-term synaptic plasticity [1418]. However, 5-HT appeared to reinstate OD plasticity in adult rats [1920]. The origin of the disparity between 5-HT regulation of long-term synaptic plasticity and OD plasticity in different age groups has been unclear. In our previous report, we also demonstrated that 5-HT suppressed long-term potentiation (LTP) in adolescent (5-week-old) rats but enhanced LTP in adult (8-week-old) rats [21]. Thus, 5-HT may have different roles in adolescent and adulthood brain. However, the mechanisms underlying the opposite regulation of LTP in the two age groups have not been addressed. Studies on the underlying mechanisms of the differential regulation of LTP may provide an insight to understand the reason for the disparity between 5-HT regulation of long-term synaptic plasticity and OD plasticity in different age groups.
Thus, in the present study, we investigated how 5-HT regulates the induction of LTP in opposite direction at different ages. To address this, we investigated the 5-HT modulation of apha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor (AMPAR)-, GABAAR-, and NMDAR-mediated synaptic transmissions and their involvement in the 5-HT regulation of LTP. Enhanced GABAAR-mediated transmission underlay the 5-HT suppression of LTP at 5 weeks. On the contrary, facilitation of LTP at 8 weeks appeared to be mediated by the enhanced function of GluN2B subunit-containing NMDAR, which could be observed only at 8 weeks. These results suggest that 5-HT could induce different metaplastic changes in the visual cortical network depending on the developmental stages.
Go to : 
METHODS
Slice preparation
Visual cortical slices were prepared from 5- (P35 to P41) and 8-week-old (P56 to P62) Sprague-Dawley rats of either sex (Orientbio Inc., Seoul, Korea), which were raised under the standard conditions (23±1℃, 12/12 hours light/dark cycle). Animal care and surgical procedures were conducted with the approval of the Institutional Animal Care and Use Committee of the School of Medicine at The Catholic University of Korea, and were consistent with the National Institutes of Health Guide for the Care and Use of Laboratory Animals. The animals were sedated with chloral hydrate (400 mg/kg, i.p.) before decapitation. The brains were quickly removed to cold dissection medium consisting of (in mM) 125 NaCl, 2.5 KCl, 1 CaCl2, 2 MgSO4, 1.25 NaH2PO4, 25 NaHCO3, and 10 D-glucose, bubbled with carbogen (95% O2/5% CO2). Then coronal slices of the occipital cortex were prepared in 300 µm of thickness on a vibrotome (Campden Instruments, Leics, UK). The slices were recovered for 40 min at 37℃ in a submerging chamber with carbogenated dissection medium, and were maintained at room temperature before recording.
Recording of field excitatory postsynaptic potential (fEPSP) and LTP induction
Recording electrodes (1~2 MΩ) were pulled from borosilicate glass pipettes (1B150F-4, World Precision Instruments, Inc., Sarasota, FL, USA) using a micropipette puller (MODEL P-97, Sutter Instrument Co., Novato, CA, USA). The recording pipette was filled with artificial cerebrospinal fluid (ACSF) and the tip of the pipette was located at layer 2/3 of the visual cortical slices. ACSF consisted of (in mM) 125 NaCl, 2.5 KCl, 2 CaCl2, 1 MgSO4, 1.25 NaH2PO4, 25 NaHCO3, and 10 D-glucose. A brief rectangular current pulse (0.2 ms) was applied to evoke fEPSP with a concentric bipolar electrode located in layer 4. Stimulus intensities were adjusted to evoke fEPSP with half maximal amplitude. After more than 10 min of stable baseline recording of fEPSP, theta-burst stimulation (TBS) was applied to induce LTP. The TBS consisted of five bursts (5 Hz) of ten pulses at 100 Hz, which was applied five times (10 s interval) with the test stimulus intensity. The peak amplitude of the fEPSP, measured at 30~40 min after TBS, was compared to the baseline responses to analyze the effect of TBS.
Whole-cell voltage clamp recording
Whole-cell patch clamp recording was conducted with an EPC8 amplifier (HEKA Elektronik, Lambrecht, Germany) and pClamp 9.0 software (Axon Instruments, Foster City, CA, USA). Slices were placed in a recording chamber containing carbogenated ACSF (1.5~2 ml/min) at 32~33℃. Pyramidal neurons in layer 2/3 of the primary visual cortex were visually identified using IR-DIC video-microscopy with an upright microscope (BX51-WI fitted with a 40×/0.80 NA water immersion objective; Olympus, Tokyo, Japan). Whole-cell configuration was achieved with the recording electrodes (3~4 MΩ), and regular spiking patterns were confirmed with the square current injection. Typical access resistance was 15~20 MΩ. Data were low-pass filtered at 5 kHz and sampled at 10 kHz. K-gluconate-based pipette solution, consisting of (in mM) 130 K-gluconate, 10 KCl, 4 Mg-ATP, 10 Na2-phosphocreatine, 0.3 Na3-GTP, and 10 HEPES (pH 7.25 with KOH), was used to record AMPAR and NMDAR currents. CsCl-based pipette solution, consisting of (in mM) 145 CsCl, 4 Mg-ATP, 10 Na2-phosphocreatine, 0.3 Na3-GTP, 10 HEPES, and 3 QX-314 (pH 7.25 by CsOH), was used to record GABAAR currents. Receptor currents were evoked at -70 mV holding potential by electrical stimulation of the underlying layer 4 with a concentric bipolar tungsten electrode. AMPAR currents were recorded with normal ACSF. The AMPAR antagonist 6,7-dinitroquinoxaline-2,3-dione (DNQX, 20 µM) and the NMDAR antagonist D-aminopentanoate (D-AP5, 50 µM) were applied to record GABAAR currents. For the recording of NMDAR currents, DNQX and the GABAAR antagonist bicuculline (10 µM) were added to low Mg2+ (0.4 mM) ACSF. Amplitude of AMPAR, GABAAR, and NMDAR currents were adjusted to -500, -500, and -100 pA, respectively, for stable recording. Effects of 5-HT or 5-HT agonists on the receptor currents were assessed after 7 min of drug application. In case of AMPAR and GABAAR, the peak amplitude was compared. However, the measurement of the amplitude of NMDAR can be ambiguous due to the presence of dual peaks especially at 5 weeks. Thus, area under the current was measured to evaluate the changes of NMDAR currents. Area was calculated from the beginning of the evoked NMDAR currents to 400 ms after. The decay phase of NMDAR current was fitted by a single exponential function to assess the decay time constant, as follows:
Chemicals
The PKA inhibitor 6~22 amide (PKI) was purchased from Calbiochem (La Jolla, CA). DNQX, D-AP5, ifenprodil, bicuculline, 2-me-5-HT, NAN-190, 8-hydroxy-N,N-dipropyl-2-aminotetralin (DPAT), and 2,5-dimethoxy-4-iodoamphetamine (DOI) were purchased from Tocris (Bristol, UK). The other chemicals were purchased from Sigma (St. Louis, MO, USA).
Statistical analysis
Data are expressed as the mean±SE. Statistical comparisons were performed using paired or unpaired two-tailed Student's t-tests. The level of significance was set at p<0.05.
Go to :
RESULTS
To identify the underlying mechanisms of the differential regulation of LTP induction between different age groups, we investigated the differences in 5-HT regulation of AMPAR-, GABAAR-, and NMDAR-mediated neurotransmissions between 5 and 8 weeks in the present study. Then we investigated if the differential regulation of neurotransmission underlies the differential regulation of LTP induction.
Opposite effects of 5-HT on LTP induction between 5 and 8 weeks
First, we reevaluated how 5-HT affects the induction of LTP at 5 and 8 weeks (Fig. 1). After stabilizing the baseline fEPSP recording at layer 2/3, TBS was applied to layer 4 to induce LTP. At 5 weeks, LTP induced by TBS (116.65±3.45%, n=9, p<0.01 vs. baseline) was inhibited by 10 µM of 5-HT (104.24±2.71%, n=9, p=0.157 vs. baseline, p<0.05 vs. control) (Fig. 1A). At 8 weeks, TBS did not induce LTP (98.97±2.05%, n=8, p=0.637 vs. baseline) but 5-HT reinstated LTP (117.38±3.47%, n=9, p<0.01 vs. baseline, p<0.001 vs. control) (Fig. 1B), which was in contrast to 5 weeks. Thus, 5-HT suppresses LTP at 5 weeks but facilitates LTP at 8 weeks. These results were consistent with our previous results [21]. The LTPs induced with control ACSF at 5 weeks and with 5-HT at 8 weeks were all inhibited by the NMDAR antagonist D-AP5 (98.21±1.51%, n=7, p=0.284 vs. baseline for 5 weeks; 95.86±2.29%, n=8, p=0.113 vs. baseline for 8 weeks), suggesting that these LTPs are NMDAR-dependent (Fig. 1C).
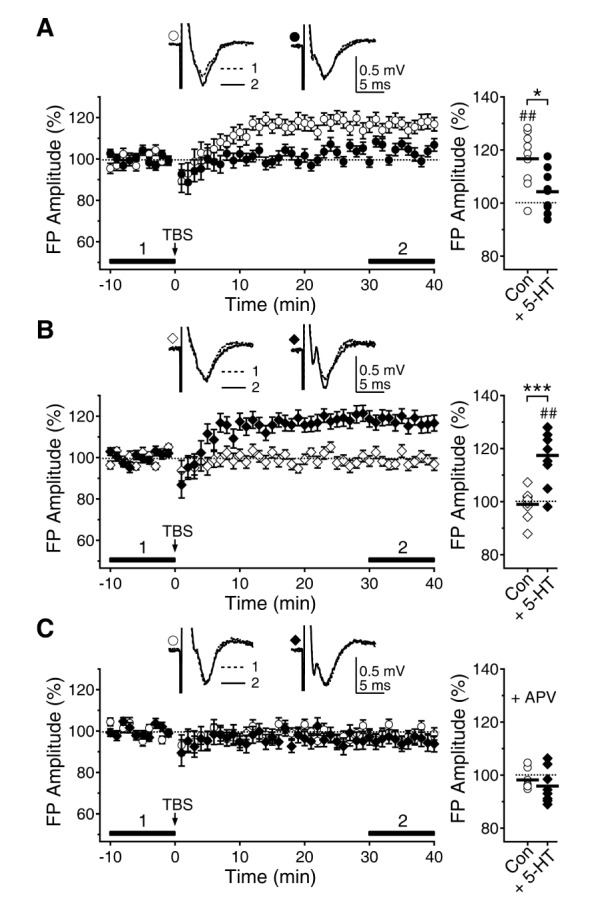 | Fig. 1Opposite effects of 5-HT on LTP induction at 5 and 8 weeks. TBS was applied to layer 4 to induce LTP of fEPSP recorded at layer 2/3. (A) LTP induced by TBS (open circle) was inhibited by 5-HT (10 µM, closed circle) at 5 weeks. Left panel plots the amplitude of fEPSPs normalized to the baseline fEPSPs. Left upper traces show average recordings taken from representative experiments at the indicated time periods. Right panel shows individual data (symbols) and averages (thick lines) of the amplitude of fEPSPs, which were measured 30 to 40 min after TBS. ##p<0.01 vs. baseline, *p<0.05 between groups linked by lines. (B) At 8 weeks, LTP could not be induced by TBS (open diamond). However, LTP was reinstated by 5-HT (closed diamond). ##p<0.01 vs. baseline, ***p<0.001 between groups linked by lines. (C) Application of the NMDAR antagonist D-AP5 (+APV) inhibited the LTPs with control ACSF at 5 weeks (open circle) and with 5-HT at 8 weeks (closed diamond).
|
Effects of 5-HT on AMPAR-, GABAAR-, and NMDAR-mediated currents
Next, we investigated how 5-HT affects AMPAR-, GABAAR-, and NMDAR-mediated neurotransmission in layer 2/3 pyramidal neurons at 5 and 8 weeks. Receptor currents were evoked by electrical stimulation of the underlying layer 4. Cells were maintained at -70 mV holding potential. AMPAR currents were recorded with K-gluconate-based pipette solution. Amplitude of AMPAR currents were not affected by 5-HT at both 5 (511.63±8.38 pA to 505.53±10.54 pA, n=10, p=0.471) and 8 weeks (494.84±10.22 pA to 498.46±12.03 pA, n=9, p=0.631) (Fig. 2A). GABAAR currents were recorded with CsCl-based pipette solution in the presence of DNQX and D-AP5. Amplitude of GABAAR currents were enhanced by 5-HT at both 5 (493.82±9.31 pA to 584.73±22.01 pA, n=11, p<0.01) and 8 weeks (486.07±10.43 pA to 538.39±14.61 pA, n=11, p<0.01) (Fig. 2B). The increase at 8 weeks (10.7%) appeared to be smaller than at 5 weeks (18.4%), but the difference was not significant (p=0.128). NMDAR currents were recorded with K-gluconate-based pipette solution in the presence of DNQX and bicuculline. Mg2+ in ACSF was lowered to 0.4 mM. Area under the current was analyzed for NMDAR currents, instead of amplitude, because dual peaks were observed in many cases at 5 weeks as shown in Fig. 2C1. However, only single peak with fast kinetics was observed at 8 weeks. In a recent study, we demonstrated that the fast and slow peaks are mediated by GluN2A and GluN2B subunit-containing NMDAR, respectively [22]. Thus, we speculate that NMDARs containing GluN2B subunit appeared to minimally participate in the synaptic NMDAR currents at 8 weeks. In adult neocortex, major proportion of GluN2B subunit-containing NMDARs is located in extrasynaptic sites [2324]. The changes in NMDAR currents kinetics between 5 and 8 weeks in the present study (Fig. 2C) may reflect the translocation of GluN2B subunit-containing NMDARs to extrasynaptic sites. NMDAR currents were not affected by 5-HT at 5 weeks (12703.35±847.26 pA·ms to 13051.95±814.77 pA·ms, n=10, p=0.168) (Fig. 2C1). However, 5-HT enhanced NMDAR currents at 8 weeks (8764.76±435.46 pA·ms to 11285.74±746.17 pA·ms, n=12, p<0.01) (Fig. 2C2). Decay time constant of NMDAR current was also increased by 5-HT at 8 weeks (52.14±1.49 ms to 73.21±4.97 ms, p<0.01). The slower decay suggests that the increase in NMDAR currents may reflect the increase in the GluN2B subunit-containing NMDAR component, since GluN2B subunit shows slower kinetics than GluN2A subunit [25]. To address this, we tried to investigate the effects of NVP-AAM077, ifenprodil, CP-101,606, and PPDA, which are known as preferential blockers of GluN2A, GluN2B, GluN2B and GluN2C/D subunits, respectively, on 5-HT-mediated enhancement of NMDAR currents at 8 weeks. In terms of selectivity, NVP-AAM077 appeared to be not suitable, since it showed significant inhibition of both GluN2A and GluN2B subunits-mediated currents at 5 weeks even at 30 nM concentration (data not shown). Thus, we investigated the effects of ifenprodil, CP-101,606, and PPDA (Fig. 3). NMDAR currents were decreased by about 24% by ifenprodil (3 µM) (7681.12±493.52 pA·ms to 5906.62±402.55 pA·ms, n=8, p<0.001) and 5-HT had no effect in the presence of ifenprodil (to 5780.12±370.41 pA·ms, p=0.099) (Fig. 3A). CP-101,606 (3 µM) decreased NMDAR currents by about 33% (7843.71±469.04 pA·ms to 5267.14±312.01 pA·ms, n=7, p<0.001) and 5-HT had no effect in the presence of CP-101,606 (to 5126.71±350.52 pA·ms, p=0.196) (Fig. 3B). PPDA (300 nM) slightly decreased NMDAR currents by about 5% (8317.42±554.42 pA·ms to 7869.86±437.15 pA·ms, n=7, p<0.05) and 5-HT enhanced the NMDAR currents in the presence of PPDA (to 9615.42±781.84 pA·ms, p<0.01) (Fig. 3C). These results indicate that the increase in NMDAR currents by 5-HT was indeed mediated by the increase in the GluN2B subunit-containing NMDAR component. Thus, the main difference in 5-HT regulation between 5 and 8 weeks was the enhancement of the GluN2B subunit-containing NMDAR at 8 weeks.
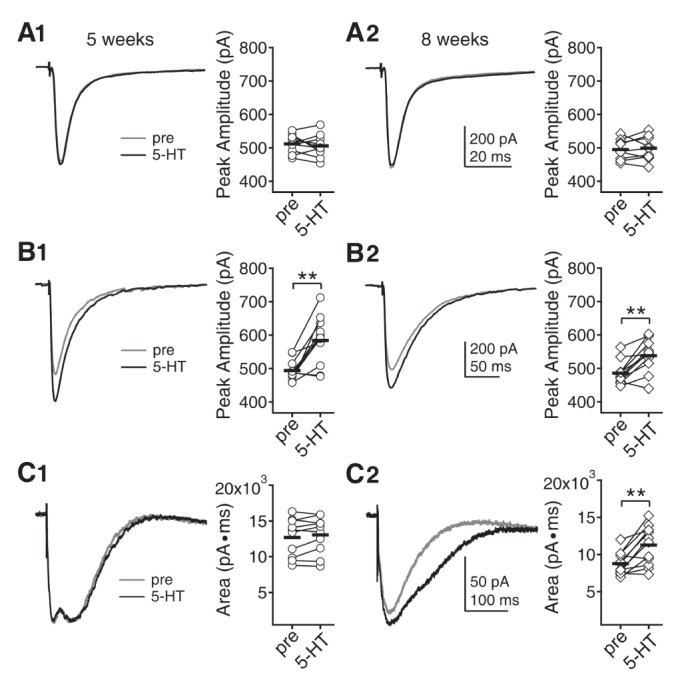 | Fig. 2Effects of 5-HT on AMPAR-, GABAAR-, and NMDAR-mediated currents in layer 2/3 pyramidal neurons at 5 and 8 weeks. (A) AMPAR currents were evoked by electrical stimulation of the underlying layer 4 and were recorded with K-gluconate-based pipette solution at -70 mV holding potential. (A1) Effect of 5-HT on AMPAR currents at 5 weeks. Left panel shows traces from a representative recording. Right panel shows individual data (symbols) and averages (thick lines) of the amplitude of AMPAR currents, which were measured before and 7 min after 5-HT application. (A2) Effect of 5-HT on AMPAR currents at 8 weeks. (B) GABAAR currents were recorded with CsCl-based pipette solution at -70 mV holding potential. Amplitude of GABAAR currents were compared before and 7 min after 5-HT application. (B1) Effect of 5-HT on GABAAR currents at 5 weeks. (B2) Effect of 5-HT on GABAAR currents at 8 weeks. **p<0.01 between groups linked by lines. (C) NMDAR currents were recorded with K-gluconate-based pipette solution at -70 mV holding potential in low Mg2+ (0.4 mM) ACSF. Area under the current was compared before and 7 min after 5-HT application. (C1) Effect of 5-HT on NMDAR currents at 5 weeks. (C2) Effect of 5-HT on NMDAR currents at 8 weeks.
|
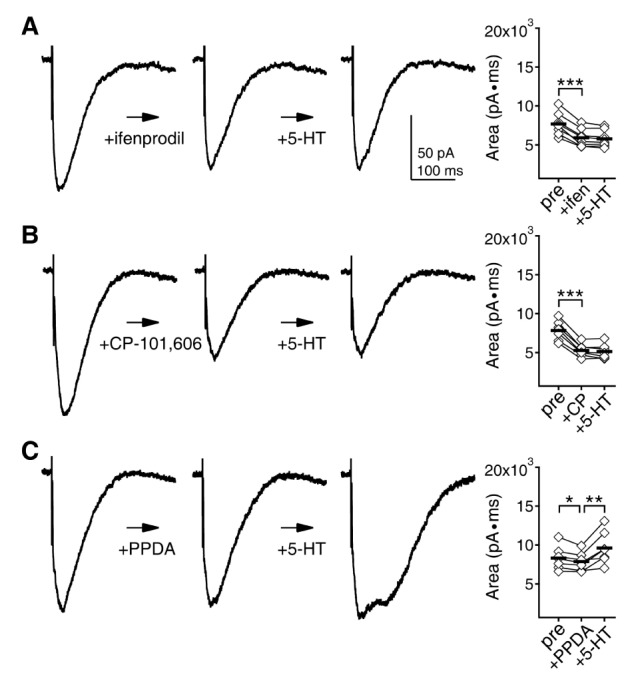 | Fig. 3Importance of GluN2B subunit in the 5-HT-mediated enhancement of NMDAR currents. After 7 min application of ifenprodil, CP-101,606, and PPDA, which are known as preferential blockers of GluN2B, GluN2B and GluN2C/D subunits, respectively, 5-HT was added to the ACSF for 7 min. (A) 5-HT had no effect on NMDAR currents in the presence of ifenprodil (3 µM). Left panel shows traces of NMDAR currents at the baseline condition, after ifenprodil application, and after 5-HT application from a representative recording. Right panel shows individual data (symbols) and averages (thick lines) of the changes in the area under the NMDAR currents. (B) 5-HT had no effect on NMDAR currents in the presence of CP-101,606 (3 µM). (C) 5-HT enhanced NMDAR currents in the presence of PPDA (300 nM). *p<0.05, **p<0.01 and ***p<0.001 between groups linked by lines.
|
In a previous report, we demonstrated that synaptic GABAAR currents are regulated by the activity of Ca2+/calmodulin-dependent protein kinase II (CaMKII) [26]. Enhancement of GABAAR currents by the activation of 5-HT2 receptor at 5 weeks was shown to be mediated by CaMKII [27]. We confirmed that 5-HT2 receptor and CaMKII are also responsible for the enhancement of GABAAR currents at 8 weeks in the present study (data not shown). The next question should be which molecular pathways are involved in the 5-HT modulation of NMDAR at 8 weeks. The 5-HT1A receptor antagonist NAN-190 (10 µM) had no effect on 5-HT enhancement of NMDAR currents (8966.51±414.08 pA·ms to 11151.68±640.23 pA·ms, n=10, p<0.01) (Fig. 4A1). However, the 5-HT2 receptor antagonist mesulergine (10 µM) blocked the effect of 5-HT on NMDAR (8525.64±417.94 pA·ms to 8838.51±480.85 pA·ms, n=11, p=0.081) (Fig. 4A2). In accordance with these results, the 5-HT1A receptor agonist DPAT (10 µM) had no effect on NMDAR currents (9062.85±424.67 pA·ms to 8986.9±410.06 pA·ms, n=9, p=0.567), but the 5-HT2 receptor agonist DOI (10 µM) enhanced NMDAR currents (8345.91±384.55 pA·ms to 10655.42±736.12 pA·ms, n=10, p<0.01) (Fig. 4B). The PKA, PKC, and tyrosine kinases inhibitors PKI (100 µ g/ml), chelerythrine (50 µM), and genistein (50 µM) in the pipette solution did not affect the 5-HT enhancement of NMDAR (8871.84±476.37 pA·ms to 11076.6±781.08 pA·ms, n=10, p<0.01 for PKI; 8753.06±598.61 pA·ms to 11251.94±996.65 pA·ms, n=9, p<0.01 for chelerythrine; 8053.2±448.63 pA·ms to 9983.52±561.01 pA·ms, n=10, p<0.01 for genistein) (Fig. 5A~C). However, the CaMKII inhibitor KN-93(10 µM) in the pipette solution blocked the effect of 5-HT on NMDAR (9055.6±590.92 pA·ms to 9242.59±622.2 pA·ms, n=10, p=0.365) (Fig. 5D). These results indicate that 5-HT enhancement of NMDAR currents at 8 weeks was mediated by 5-HT2 receptor and CaMKII, which is the same as the 5-HT enhancement of GABAAR.
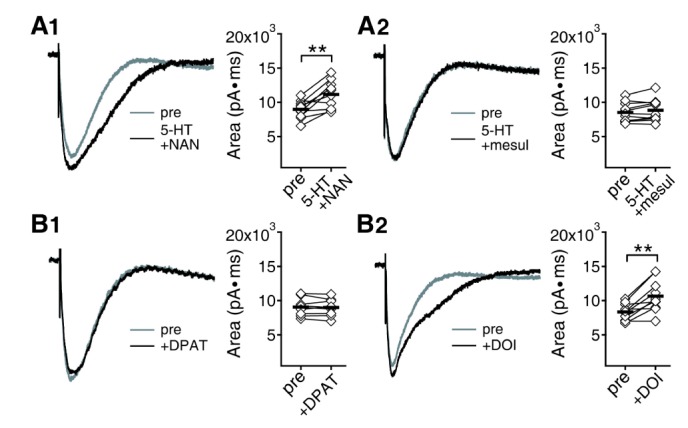 | Fig. 4Effects of 5-HT receptor antagonists on 5-HT enhancement of NMDAR currents and 5-HT receptor agonists on NMDAR currents at 8 weeks. (A) The 5-HT1A receptor antagonist NAN-190 (A1) or the 5-HT2 receptor antagonist mesulergine (A2) were co-applied with 5-HT. Area under the current was compared before and 7 min after the application of each drug combination. Left panel shows traces from a representative recording. Right panel shows individual data (symbols) and averages (thick lines) of the changes in area under the current. **p<0.01 between groups linked by lines. (B) Effects of the 5-HT1A receptor agonist DPAT (B1) or the 5-HT2 receptor agonist DOI (B2) on NMDAR currents.
|
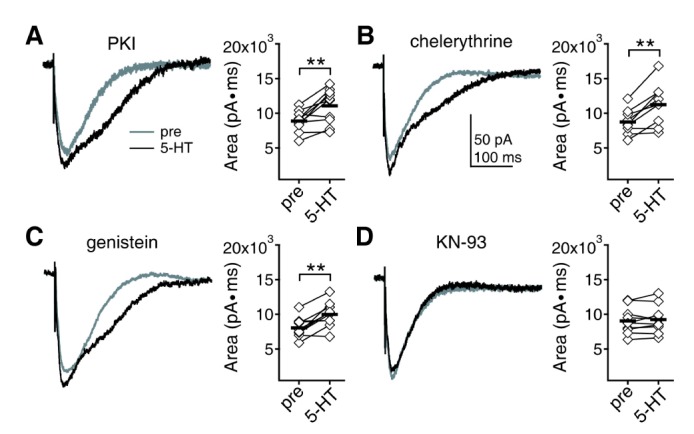 | Fig. 5Effects of kinases inhibitors on 5-HT enhancement of NMDAR currents at 8 weeks. (A) 5-HT was applied in the presence of the PKA inhibitor PKI in pipette. Area under the current was compared before and 7 min after the application of 5-HT. Left panel shows traces from a representative recording. Right panel shows individual data (symbols) and averages (thick lines) of the changes in area under the current. **p<0.01 between groups linked by lines. (B) 5-HT was applied in the presence of the PKC inhibitor chelerythrine in pipette. (C) 5-HT was applied in the presence of the tyrosine kinases inhibitor genistein in pipette. (D) 5-HT was applied in the presence of the CaMKII inhibitor KN-93 in pipette.
|
Reversal of 5-HT effects by GABAAR antagonist and GluN2B subunit selective NMDAR antagonist
Results from previous sections demonstrated that 5-HT selectively enhanced GABAAR-mediated neurotransmission at 5 weeks. In 8 weeks, both GABAAR- and NMDAR-mediated neurotransmissions were enhanced by 5-HT via 5-HT2 receptor and CaMKII. If these 5-HT modulations of neurotransmission were responsible for the opposite regulation of LTP at 5 and 8 weeks, manipulations which can compensate the changes in neurotransmission may reverse the effects of 5-HT on LTP. To address this, we assessed the concentration of the GABAAR antagonist bicuculline and the GluN2B subunit selective NMDAR antagonist ifenprodil to reverse the increase in GABAAR- and NMDAR-mediated currents by 5-HT. Bicuculline and ifenprodil were applied 7 min after the application of 5-HT (Fig. 6). At 300 nM, bicuculline reversed the 5-HT enhancement of GABAAR-mediated currents (511.97±10.34 pA to 598.11±18.32 pA by 5-HT, to 501.05±16.79 pA by bicuculline, n=11, p=0.587 between control and bicuculline) (Fig. 6A). Ifenprodil reversed the 5-HT effects on NMDAR at 1 µM (8851.05±569.46 pA·ms to 11001.46±728.18 pA·ms by 5-HT, to 8959.67±580.92 pA·ms by ifenprodil, n=12, p=0.659 between control and ifenprodil) (Fig. 6B).
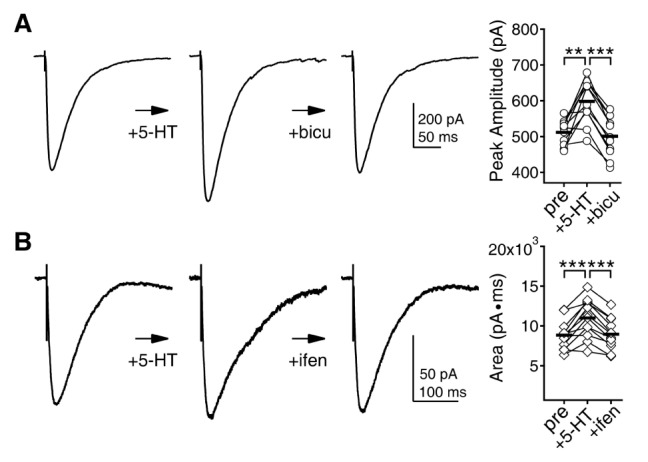 | Fig. 6Effects of bicuculline and ifenprodil on 5-HT modulation of GABAAR and NMDAR currents. The effects of 5-HT on GABAAR and NMDAR currents at 5 and 8 weeks, respectively, were assessed first, and then bicuculline and ifenprodil were applied. (A) Bicuculline (300 nM) can negate the effect of 5-HT on GABAAR currents at 5 weeks. Left panel shows traces of GABAAR currents at the baseline condition, after 5-HT application, and after bicuculline application from a representative recording. Right panel shows individual data (symbols) and averages (thick lines) of the changes in the amplitude of GABAAR currents. **p<0.01 and ***p<0.001 between groups linked by lines. (B) Ifenprodil (1 µM) can negate the effect of 5-HT on NMDAR currents at 8 weeks. Left panel shows traces of NMDAR currents at the baseline condition, after 5-HT application, and after ifenprodil application from a representative recording. Right panel shows individual data (symbols) and averages (thick lines) of the changes in the area under the NMDAR currents.
|
Now, we can negate the effects of 5-HT on GABAAR currents at 5 weeks and NMDAR currents at 8 weeks with 300 nM of bicuculline and 1 µM of ifenprodil, respectively. We investigated the effects of 300 nM of bicuculline and 1 µM of ifenprodil on 5-HT suppression of LTP at 5 weeks and 5-HT facilitation of LTP at 8 weeks (Fig. 7). Bicuculline reinstated the 5-HT suppressed LTP at 5 weeks (118.89±2.78%, n=9, p<0.001 vs. baseline) (Fig. 7A). Application of both bicuculline and ifenprodil reversed the effect of 5-HT on LTP at 8 weeks (103.21±2.26%, n=10, p=0.162 vs. baseline) (Fig. 7B). In addition, ifenprodil alone was enough to inhibit the 5-HT facilitated LTP at 8 weeks (101.58±2.17%, n=9, p=0.487 vs. baseline). These results collectively suggest that 5-HT suppresses LTP at 5 weeks by enhancing GABAAR-mediated neurotransmission and facilitates LTP at 8 weeks by enhancing GluN2B subunit-containing NMDAR. Thus, the opposite regulation of LTP by 5-HT originates from its differential regulation of neurotransmission between different age groups.
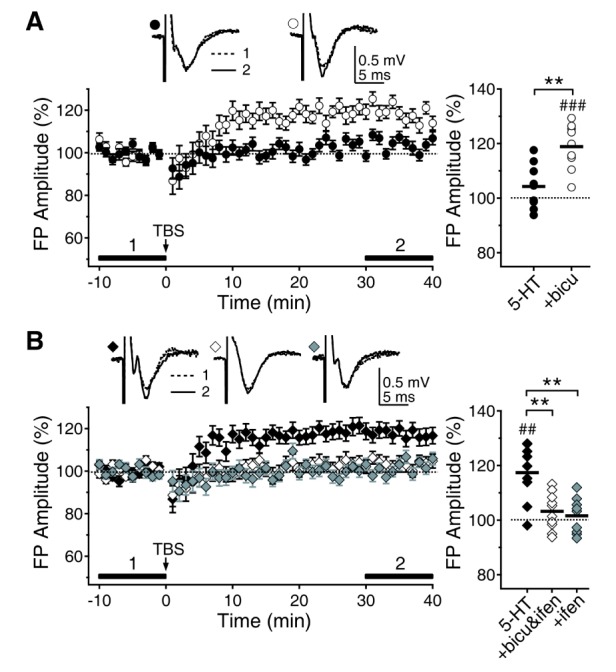 | Fig. 7Reversal of the 5-HT regulation of LTP by bicuculline and ifenprodil. For clear comparison, data for 5-HT were reproduced from Fig. 1. (A) 5-HT suppression of LTP at 5 weeks (closed circle) was rescued by the application of bicuculline (300 nM) (open circle). (B) 5-HT enhancement of LTP at 8 weeks (closed diamond) was vanished by the application of both bicuculline and ifenprodil (1 µM, open diamond) or ifenprodil alone (grey diamond). ##p<0.01, ###p<0.001 vs. baseline, **p<0.01 between groups linked by lines.
|
Go to :
DISCUSSION
In the present study, we investigated the differences in 5-HT regulation of AMPAR-, GABAAR-, and NMDAR-mediated neurotransmission between the visual cortical slices of 5 and 8 week-old-rats and their roles in the regulation of LTP, to explain how 5-HT differentially regulates LTP between 5 and 8 weeks. Enhanced GABAAR currents underlay the 5-HT suppression of LTP at 5 weeks. Enhancement of GluN2B subunit-containing NMDAR, which could only be observed at 8 weeks, appeared to facilitate the induction of LTP at 8 weeks. Thus, the differential regulation of neurotransmission may underlie the differential regulation of LTP by 5-HT between the age groups.
Regulation of neurotransmission by 5-HT
Excitation-inhibition balance in cortical network can be regulated by 5-HT, since excitatory and inhibitory neurotransmissions can be differentially regulated by 5-HT [151628]. AMPAR-, GABAAR-, and NMDAR-mediated transmissions are all subject to 5-HT regulation. In hippocampus, 5-HT suppressed AMPAR-mediated component via 5-HT1A receptor [29]. 5-HT potentiated GABAAR-mediated currents in rat sacral dorsal commissural neurons via 5-HT2 receptor and PKC [30]. In the visual cortex, 5-HT enhancement of GABAAR currents was mediated by 5-HT2 receptor and CaMKII [27]. On the contrary, the activation of 5-HT2 receptor suppressed GABAAR currents in the prefrontal cortex [31]. Since GABAAR-mediated neurotransmission critically regulates the activation of NMDAR [3233], 5-HT may be able to regulate the NMDAR-dependent plasticity by modulating GABAAR currents. Furthermore, 5-HT can more directly regulate NMDAR-dependent plasticity by modulating NMDAR currents. Suppression of NMDAR component was suggested to underlie the inhibition of LTP by 5-HT in the visual cortex [34]. Increased NMDAR function by 5-HT2 receptor in basolateral amygdala appeared to facilitate LTP induction [35]. In addition, 5-HT can selectively regulate NMDAR subunits. GluN2B subunit can be selectively removed from the synapses by the activation of 5-HT1A receptor in the prefrontal cortex [36]. 5-HT2 receptor appeared to counteract the effect of 5-HT1A receptor on GluN2B subunit [37]. In this case, CaMKII was involved in the decrease in GluN2B subunit component. In our present study, CaMKII underlay the 5-HT2 receptor-mediated increase in GluN2B subunit component. These results demonstrate that 5-HT modulation of neurotransmission can be very different in various regions of the brain. This diversity may originate from the differences in the distribution of 5-HT receptor subtypes in different regions of the brain [38] and the diverse biochemical pathways associated with various 5-HT receptor subtypes [39]. Age can add more complexity as shown by the differential regulation of NMDAR currents between age groups in the present study. We speculate that the downstream biochemical pathways associated with a 5-HT receptor subtype can change or expand their connections to various molecular targets through development, since 5-HT2 receptor regulates only the GABAAR currents at 5 weeks but regulates both GABAAR and NMDAR currents at 8 weeks. GluN2B subunit-containing NMDARs are thought to exist in extrasynaptic sites at 8 weeks. We speculate that 5-HT2 receptor and CaMKII selectively translocate extrasynaptic NMDARs to synaptic sites at 8 weeks but do not affect synaptic NMDARs at 5 weeks. This assumption could possibly explain the 5-HT-induced increase in GluN2B subunit component at 8 weeks. Furthermore, the developmental changes in the expression patterns of 5-HT receptor subunits may contribute to the differential effects of 5-HT between different ages [4041]. However, this may not be a plausible explanation for the differences between 5 and 8 weeks, since the expression level of 5-HT2 receptors does not change much after early developmental period in neocortex [4142].
Metaplastic regulation of long-term synaptic plasticity by 5-HT
Metaplasticity is defined as a change in the ability to induce synaptic plasticity [43]. In the present study, 5-HT changed the inducibility of LTP at 5 and 8 weeks. Thus, the visual cortical network appears to be under the metaplastic control of 5-HT. There have been some confusing reports on 5-HT regulation of plasticity in the visual cortex. In kitten visual cortex, 5-HT is thought to be necessary for OD plasticity [1744] and long-term synaptic plasticity can be facilitated by 5-HT [45]. However, long-term synaptic plasticity was suppressed by 5-HT in the visual cortex of juvenile rats [144647]. In the more aged rats, 5-HT rather enhanced long-term synaptic plasticity [21]. The different metaplastic control by 5-HT, at different age groups in the present study, could provide important insight to understand this disparity. 5-HT may control the induction of plasticity differentially depending on the ages and species, because it can show tremendous amount of variability in the regulation of functions of various neurotransmitter receptors.
Metaplasticity has been generally thought to be induced by prior activities in the neural network [48]. The changes in the GABAAR-mediated neurotransmission can induce metaplasticity, if the activities in postsynaptic neurons were enough to send retrograde endocannabinoid-mediated signal to presynaptic GABAergic terminals [49]. Activities also regulate the expression of NMDAR subunits in the visual cortex [5051]. Activity-dependent changes in the GluN2A/GluN2B ratio appeared to be involved in the metaplasticity [95253]. In the present study, 5-HT was shown to regulate GABAAR-mediated transmission and GluN2B subunit-containing NMDAR component. These effects of 5-HT appeared to underlie the changes in the inducibility of LTP. These results indicate that neuromodulatory regulation of GABAAR- and NMDAR-mediated neurotransmissions can induce metaplasticity in the visual cortex. Thus, not only the activity-induced metaplasticity but also the neuromodulator-induced metaplasticity should be considered to fully understand the metaplastic control of neural network.
Enhancing synaptic plasticity in adult cortical network
Since synaptic plasticity is critical in the network formation and stabilization [54], the mechanisms behind the developmental decline of plasticity in the visual cortex have been of interest [15556]. Efforts to restore the plasticity in adult followed as a necessity. To this end, the roles of extracellular matrix, GABAergic inhibition, and NMDAR have been extensively studied. Extracellular matrix is an important regulator of dendritic spine dynamics and visual cortical plasticity [8]. After full development, the extracellular matrix stabilizes the neural connectivity and decreases synaptic plasticity. Thus, synaptic plasticity could be restored in adult brain when extracellular matrix was degraded [11]. GABAergic inhibition is also an important target for restoring plasticity in adult brain [57]. Manipulations which reinstate plasticity accompanied the reduction in GABAergic inhibition [102058]. The reduction of cortical inhibition itself was enough to promote plasticity in adult brain [59]. The juvenile composition of NMDAR subunits with higher GluN2B subunit-containing NMDAR appeared to be involved in the restoration of plasticity in the adult visual cortex [952]. Thus, many factors are involved in the developmental decline of plasticity and the restoration of plasticity in adult brain. Furthermore, we should also consider the regulation of plasticity by neuromodulators [606162]. One of the particularly interesting neuromodulators is 5-HT. The selective serotonin reuptake inhibitor fluoxetine has been known to restore plasticity in the adult visual cortex [19]. This effect appeared to be mediated by rather a long-term effect of 5-HT on cortical network via epigenetic remodeling of chromatin structure, which might result in increased brain-derived neurotrophic factor (BDNF) and decreased intracortical inhibition [1963]. In this case, decreased inhibition might be a critical factor for the reinstation of plasticity. In the present study, we showed that 5-HT can facilitate plasticity by enhancing GluN2B subunit-containing NMDAR component or suppress plasticity by enhancing GABAAR-mediated currents. These acute regulatory mechanisms may provide an important insight for the development of new pharmacological agents for restoring the synaptic plasticity in adult, i.e., acute and chronic effects should all be taken into account when considering 5-HT related agents.
It is now generally accepted that plasticity can be enhanced in adult nervous system by either pharmacological treatments or manipulations which shift the environmental stimulation levels [10]. Further efforts to understand the confining factors for plasticity and to facilitate plasticity in adult brain will help to make a breakthrough for the treatment of various neurodevelopmental diseases [1264].
Go to :
 XML Download
XML Download