

 PDF
PDF ePub
ePub Citation
Citation Print
Print


ABBREVIATIONS
Mib
Mind bomb
RING domain
Really Interesting New Gene domain
WT
wild type
KO
knock-out
LTP
long-term potentiation
E-LTP
early-phase LTP
L-LTP
late-phase LTP
LTD
long-term depression
NMDAR
N-methyl-D-aspartate receptor
LFS
low frequency stimulation
TBS
theta burst stimulation
HFS
high frequency stimulation
GluN2B
GluRepsilon2
pGluN2B
phosphorylated GluRepsilon2
INTRODUCTION
The Notch signaling pathway is an intracellular signal transduction mechanism, which is highly conserved throughout multicellular eukaryotic species [1]. To date, five Notch ligands and four Notch receptors have been identified in mammalian cells. As these are all type 1 transmembrane proteins, Notch signaling can occur only between physically adjacent cells [2]. During the development of invertebrate and vertebrate species, Notch signaling plays a critical role in regulating cell fate and integrating other developmental cues [3456]. During the development of the mammalian central nervous system (CNS), Notch signaling maintains the progenitor cell population and regulates its maturation [7]. Besides the well-known developmental role of Notch signaling, recent studies report that Notch signaling in the CNS is also involved in synaptic plasticity, learning, and memory [7891011]. These studies show that LTP and LTD were both impaired in hippocampal slices from transgenic mice with reduced Notch level, suggesting that Notch signaling is involved in modification of CA 1 synapses during stimulation [11]. Not only Notch 1 itself but also its ligand Jagged 1 were found at the synapse and activated by neuronal activity. When Notch 1 was conditionally deleted in postnatal hippocampus, LTP and LTD were impaired, leading to learning and short-term memory deficits [12].
The mib gene was originally found in zebrafish; it encodes the protein Mind bomb (Mib), which mediates Notch signaling by ubiquitinating the Notch ligand Delta and promoting its endocytosis [13]. Kong and his colleagues identified the mouse homologue of mib, and created knock-out (KO) transgenic mouse lines for Mind bomb-1 (Mib1) and its paralogue Mind bomb-2 (Mib2) [1415]. The E3 ubiquitin ligase activity of both Mib1 and Mib2 is mediated by the C-terminal Really Interesting New Gene (RING) domain, and Mib1 and Mib2 interact with Xenopus Delta (XD) via their N-terminal region [13]. When expression levels are compared in mice, Mib2 is highly expressed in adult tissues and is less abundant in embryos; however, Mib1 is highly expressed in both adult and embryotic tissues [16]. Even though zebrafish Mib and Mib2, which are orthologous to mouse Mib1 and Mib2, have redundant roles in zebrafish development [17], the different expression patterns of Mib1 and Mib2 suggest that mouse Mib1 and Mib2 may have different roles.
Even though the role of Notch signaling in development and synaptic plasticity is already known [12], the role of Mib2 in learning and memory is still unclear. In our previous study, we reported that conditional knockout of Mib1 in mature neurons of the mouse forebrain result in impaired synaptic plasticity, learning, and memory [18]. To assess whether Mib2 has a different function compared with Mib1, we used Mib2 KO mice and examined their learning and memory functions. We found impaired hippocampus-dependent long-term memory, such as contextual fear memory, in the Mib2 KO mice compared with their WT littermates. Similarly, acute hippocampal slices prepared from Mib2 KO mice exhibited impairments in various forms of LTP. When we measured protein levels in the hippocampus of these mice, we found that the level of cleaved Notch1 was lower in Mib2 KO mice than in their WT littermates after mild foot shock. These results suggest that Mib2-mediated Notch signaling is essential for regulating synaptic plasticity and memory formation in the hippocampus.
METHODS
Mice
We used 8~15-week-old male Mib2 KO and wild type (WT) littermates on the C57BL/6N genetic background for behavioral experiments. Mice were kept on a 12-h light: dark cycle, and behavioral experiments were performed during the light phase. Food and water were provided ad libitum. The Animal Care and Use Committee of Seoul National University approved the animal protocols used in this study.
Behavioral tests
For all behavioral tasks, 8~15 weeks old male mice were used. Before performing the task, mice were placed on a shelf for at least 40 minutes for accommodation.
Morris water maze task
Mice were handled for 3 minutes 7 consecutive days before the task. After the handling, mice were put into a gray opaque cylinder-shaped tank (140 cm diameter, 100 cm height) in a room with multiple spatial cues including a water tap and a computer desk, where the experimenter sat. The tank was divided into 4 virtual quadrants, and a Perspex platform (10 cm diameter) was placed at the center of a quadrant (TQ). The tank was filled with water (20~22℃), until the water level was 1 cm higher than the platform, and white paint was added to make the water opaque. Before the 1st trial on training day 1, mice were placed on the platform for 30 seconds. On the training days, mice were released at the edge of the maze facing towards the inner wall of the tank, and were trained to reach the platform. If mice were not able to reach the platform in 60 seconds, they were taken out of the maze. The releasing point was chosen randomly in every trial. When the mice failed to reach the platform, they were guided to or placed on the platform, and were rescued from the maze after 10 seconds. When the mouse successfully reached the platform, and stayed on it for more than 1 second, it was rescued from the maze after 10 seconds. Each mouse performed 4 trials per training day. The interval between trials 1 and 2 or trials 3 and 4 was 1 minute, and the interval between trials 2 and 3 was 30~45 minutes. Each mouse performed 4 training trials per day for 5 consecutive days.
Contextual fear conditioning
Prior to contextual fear conditioning, we handled each mouse for 3 minutes on 4 consecutive days. For contextual fear conditioning, the mice were placed in a chamber for 3 minutes. After 148 seconds, they were presented with an unexpected foot shock for 2 seconds (0.4 mA). Mice were returned to the chamber for testing their fear memories after 24 hours. Freezing (immobile posture except for respiration) level was measured automatically by a computer program (FreezeFrame; Coulbourn).
Elevated plus maze task
During the test, mice were placed at the center of the plus maze, and their movement was tracked for 5 minutes by a tracking program (EthoVision 3.1; Nodulus) under fluorescent light. The maze was made of white Plexiglas, and its arms were 150-cm long. The two opposing closed arms had additional walls with a height of 20 cm.
Open field task
Mice were put in a square opaque box (40×40×40 cm). The tasks were performed under dim light, and mice were tracked with a tracking program (EthoVision 3.1; Nodulus) for 10 minutes.
Electrophysiology
Field excitatory postsynaptic potential recordings were performed as described previously [19]. Using a manual tissue chopper, transverse hippocampal slices (400-µm thick) were prepared from 4~5-week-old mice for measuring the N-methyl-D-aspartate receptor (NMDAR) mediated long-term depression (LTD) or from 8~12-week-old mice (male and female) for other protocols. Animals were deeply anesthetized with isoflurane (Hana Medical), decapitated, and their brains were removed and sectioned. Hippocampal slices were maintained in an interface chamber at 28℃ oxygenated with 95% O2 and 5% CO2 and perfused (1~1.5 ml/min) with artificial cerebrospinal fluid (ACSF; 124 mM NaCl, 2.5 mM KCl, 1 mM NaH2PO4, 25 mM NaHCO3, 10 mM glucose, 2 mM CaCl2, 2 mM MgSO4). Slices were incubated in the interface chamber for at least 2 hours. After this recovery period, extracellular field excitatory postsynaptic potentials (fEPSPs) were recorded from the Schaffer collaterals (SC) of the CA1 region using a glass pipette electrode filled with ACSF (1 MΩ). The SC were stimulated every 30 seconds using concentric bipolar electrodes (MCE-100; Kopf Instruments) placed at the CA1 region. Field potentials were amplified, low-pass filtered (GeneClamp 500; Axon Instruments), and then digitized (NI PCI-6221; National Instruments) for measurement. Data were monitored, analyzed online, and reanalyzed offline using the WinLTP program (WinLTP Ltd., winltp.com, The University of Bristol, UK). For measuring long-term potentiation (LTP) and LTD, stimulation intensity was adjusted to produce a fEPSP slope that is approximately 40% of the slice's maximum slope. Two successive responses elicited twice per minute at this test stimulation intensity were averaged, and then expressed relative to the normalized baseline. After a stable baseline was recorded, we induced LTP by high-frequency stimulation (HFS; 100 Hz stimulation, 1 second, test intensity), LTD by low-frequency stimulation (LFS; 900 stimuli, 1 Hz, test intensity), and LTP by theta burst stimulation (TBS; 3×TBS, 1 second each, test intensity). Data were monitored, analyzed online, and reanalyzed offline using the WinLTP program.
Western blot
For western blot analysis, hippocampi of 8~12 weeks old mice were used. Mice were deeply anesthetized with isoflurane and decapitated; then, their hippocampi were removed and frozen immediately in liquid nitrogen and stored at -80℃ until lysis with a lysis buffer (1.6% SDS, 10 mM TrisCl; pH 6.8). Protease inhibitor cocktail (Roche 11873580001) and protein phosphatase inhibitor cocktail (Roche 04906845001) were also added to the lysis buffer. Hippocampal tissue was lysed in 400 µl lysis buffer with the QIAGEN TissueLyser LT (50 Hz, 3 min). For analysis of cleaved Notch1, mice were given a mild foot shock (0.5 mA, 2 sec) using the same protocol as for contextual fear conditioning. Fifteen micrograms of the protein samples were run by electrophoresis, and the blotted membranes were immunolabeled with antibodies against phospho-GluRepsilon2 (pGluN2B, Sigma-Aldrich, M2442), GluRepsilon2 (GluN2B, Neuromab, 75-097), Actin (Sigma-Aldrich, A2066), Tubulin (NeuroMab 75-330), cleaved Notch1 (CST, 2421) and Mib2 (Abnova H00142678-M01A). Mib1 antibody was gift from Dr. P. Gallagher.
RESULTS
Hippocampus-dependent memory was abnormal in Mib2 KO mice
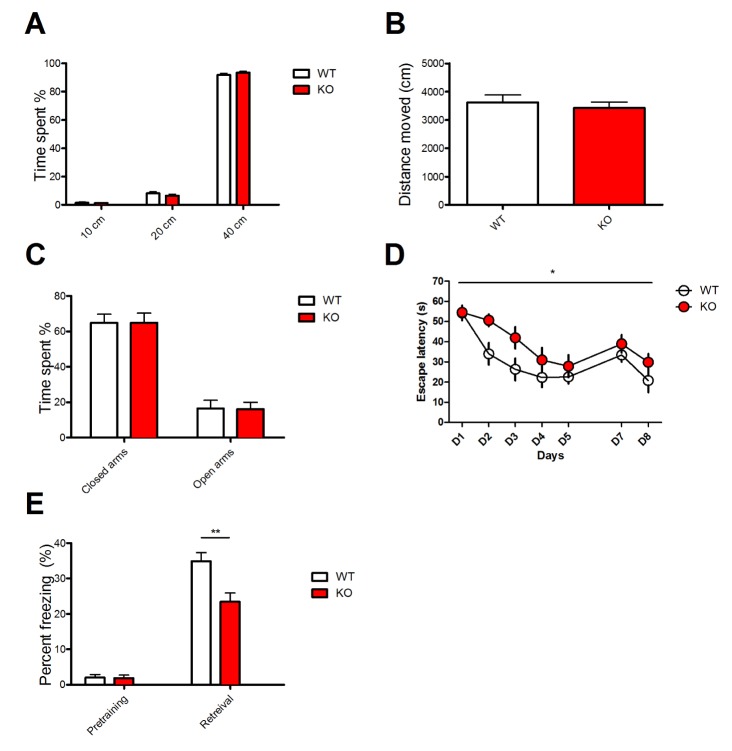
Before examining whether Mib2 deletion affects learning and memory, we tested if Mib2 KO mice exhibit abnormal anxiety. In the open field and elevated plus maze tasks, locomotive activity and anxiety level were comparable between Mib2 KO and WT mice (Fig. 1A~C). Then we tested whether mice have an intact hippocampal function using the Morris water maze [20]. In this task, mice have to learn and remember the location of a hidden platform beneath the water surface using spatial cues in the room. In the training trials, the latency to reach the hidden platform was significantly longer in the KOs than in their WT littermates (Fig. 1D). These results suggest that deletion of Mib2 impairs spatial learning in mice.
Next, we examined the mice using contextual fear conditioning test, which is another hippocampus-dependent memory task. In the training session, mice were placed in a conditioning chamber, where mild foot shocks were delivered. Then, the trained mice were exposed to the same conditioning chamber to assess their freezing behavior 24 hours after the training. Consistent with the result of the Morris water maze, Mib2 KO mice exhibited significantly less freezing behavior than WT mice (Fig. 1E), demonstrating that hippocampal learning and memory are impaired in Mib2 KO mice.
Basal synaptic transmission and LTD was normal in Mib2 KO mice
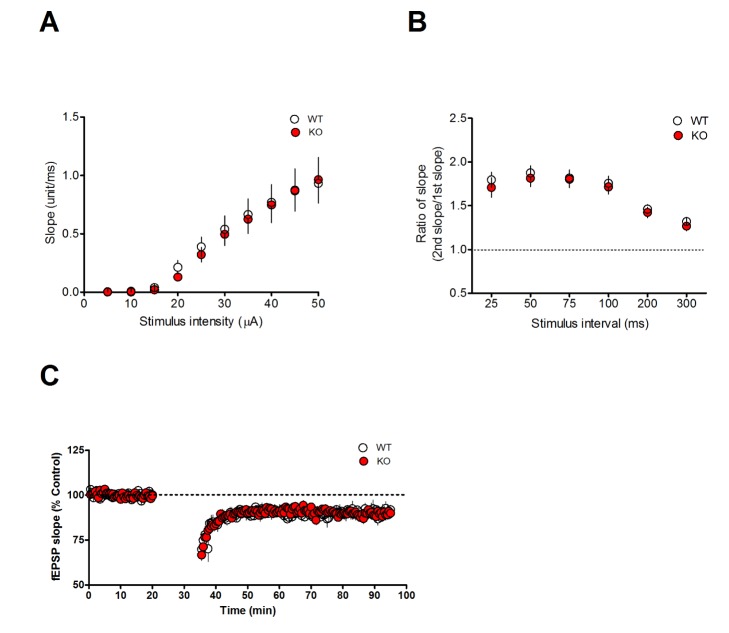
To identify the mechanism responsible for the memory deficits caused by Mib2 deletion, we examined the electrophysiological properties of Mib2 KO mice with extracellular field recordings at the SC-CA1 synapses in acute hippocampal slices. The input-output relationship and paired-pulse ratio were indistinguishable in WT and Mib2 KO mice (Fig. 2A and 2B), demonstrating that the basal synaptic transmission is intact in Mib2 KO mice. Since spatial memory of Mib2 KO mice was impaired in the NMDAR-dependent [21] water maze task, we examined the NMDAR-dependent LTD at the SC-CA1 pathway by delivering LFS (900 pulses at 1 Hz) using acute hippocampal slices. We found no significant difference in the level of LTD between genotypes (Fig. 2C), indicating that the genetic deletion of Mib2 does not affect NMDAR-LTD.
Mib2 KO mice had stimulation protocol dependent deficits in E-LTP and L-LTP
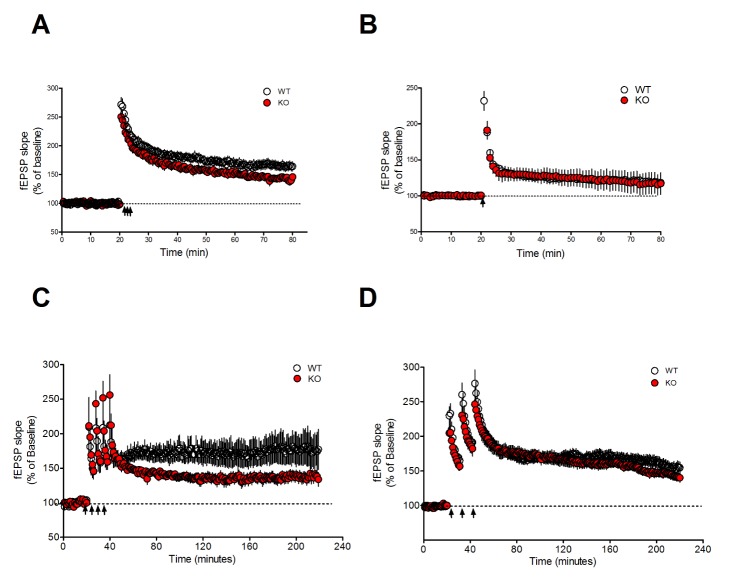
LTP at the SC-CA1 synapses plays an important role in spatial learning and memory. Mutant mice with impaired LTP often exhibit deficits in the hippocampus-dependent learning and memory [9]. Consistent with the results of behavioral tasks, which investigated the hippocampus-dependent memory, Mib2 KO mice had an abnormal TBS-induced early-phase LTP (E-LTP) (Fig. 3A). In contrast, the potentiation level of HFS (100 Hz) induced E-LTP was normal in Mib2 KO mice compared to WT controls in the last 5 minutes (Fig. 3B).
Interestingly, this protocol-dependent deficit of synaptic plasticity in Mib2 KO mice reversed during the late-phase LTP (L-LTP), which is a de novo protein synthesis dependent form of synaptic plasticity [222324]. When we induced L-LTP in the hippocampal slices by delivering four pulses of high frequency tetanus with 5 minutes intervals, L-LTP was significantly decreased in Mib2 KO mice compared to their WT littermates (Fig. 3C). However, when we induced L-LTP by delivering TBS three times with 10 minutes intervals, the potentiation level during the last 5 minutes was similar in theMib2 KO and WT littermates (Fig. 3D). These results suggest that Mib2 selectively regulates E-LTP and protein synthesis-dependent L-LTP at hippocampal SC-CA1 synapses.
These protocol-dependent deficits of synaptic plasticity in Mib2 KO mice may be explained by the difference between the two stimulation protocols. TBS and HFS both induce LTP, but they resemble two separate prominent rhythms in the brain: theta rhythm of 5 to 7 Hz and high frequency gamma rhythm of 50 to 100 Hz. While TBS is known to more accurately replicate the stimulus pattern of the naturally occurring rhythmic activity in hippocampus in vivo [25], increasing evidences show that they do not share intracellular mechanisms when inducing LTP. TBS-induced LTP requires ERK MAPK activity [26] while HFS-induced LTP does not [27]. A more recent study compared the two protocols and showed that although they share mechanisms like actin polymerization, TBS-induced LTP involves calpain-1 activation and suprachiasmatic nucleus circadian oscillatory protein degradation, while HFS-induced LTP does not. Instead, HFS requires adenosine A2 receptors and PKA [28]. TBS and HFS may represent different physiological conditions of learning and some cases have been reported where transgenic mice show LTP deficits in only one protocol [29]. Therefore, the protocol-dependent LTP and L-LTP deficit in Mib2 KO mice suggests that Mib2 is important in selective synaptic plasticity pathways.
Mib2 KO mice had a normal GluN2B level, but a diminished Notch signaling
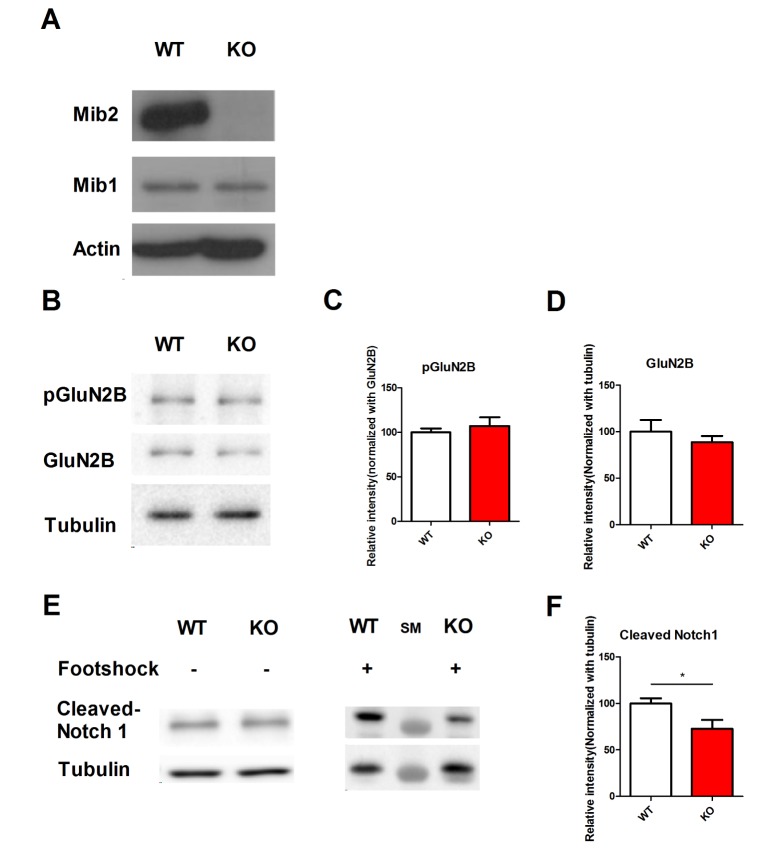
What is the molecular mechanism that caused memory impairments and synaptic plasticity deficits in Mib2 KO mice? To answer this question, we prepared hippocampal lysates from Mib2 KO and WT littermates. In the hippocampus of Mib2 KO mice, compared to the hippocampi of their WT littermates, Mib2, but not Mib1 was specifically deleted (Fig. 4A). A previous study reported that Mib2 ubiquitinates the GluN2B subunit of the NMDAR receptor [30]. However, we found no difference in the levels of pGluN2B and GluN2B between Mib2 KO and WT littermates (Fig. 4B~D). Next, we measured the level of cleaved Notch1 in the hippocampi of Mib2 KO and WT mice without foot shock and 1 hour after mild foot shock (Fig. 4E). Mib2 KO mice had a reduced level of hippocampal cleaved Notch1 after mild foot shock compared with their WT littermates (Fig. 4F). These results show that the deficits in Mib2 KO mice might be a result of impaired activity-dependent Notch signaling.
DISCUSSION
In the early developmental stages of Drosophila and zebrafish, Mib2 mediates Notch signaling by ubiquitinating the Notch ligand Delta [16]. Despite its abundant expression, the function of Mib2 in the adult mouse brain was previously unknown. In this study, we report that Mib2 plays an important role in hippocampus-dependent memory formation and synaptic plasticity by regulating Notch signaling.
Previous studies suggested that Notch signaling is involved in learning and memory formation [81031], synaptic plasticity such as LTP and LTD [11], and neuronal activity-dependent immediate early gene expression [12]. Notch1 null heterozygous knockout and conditional knockout mice exhibit normal locomotion, but impaired spatial memory [1231]. Similarly, our study showed impaired spatial learning of Mib2 KO mice in the Morris water maze test.
Are these defects in the hippocampus-dependent behavioral tasks and synaptic plasticity of Mib2 KO mice caused by impaired Notch signaling in the hippocampus? The basal level of cleaved Notch1 was similar in WT and Mib2 KO littermates, suggesting that Mib2 does not mediate basal Notch signaling. However, Mib2 KO mice had a lower level of cleaved Notch1 compared to WT mice 1 hour after a mild foot shock. Considering that hippocampal neuronal activity and synaptic plasticity are critical for contextual fear memory formation [32], Mib2 might mediate Notch signaling in the adult hippocampus during hippocampus-dependent learning and neuronal activity.
The input-output curve and paired pulse ratio confirmed that the basal transmission is not impaired in Mib2 KO mice. However, Mib2 KO mice clearly showed deficits in two different forms of LTP in a stimulation protocol dependent manner. As Notch is a transcription co-activator, and Mib2 ubiquitinates a Notch ligand [16], the deficit in L-LTP in Mib2 KO mice suggests that Mib2 might regulate L-LTP via transcription. Additionally, the deficient E-LTP in Mib2 KO mice suggests that potentiation of glutamate receptor responses might be disrupted in synapses occurring in these mice [33]. Therefore, Mib2 might be important in the formation and maintenance of synaptic plasticity.
Mib2 has been reported to ubiquitinate the GluN2B subunit of the NMDAR receptor in the postsynaptic site [30]. Therefore, we investigated if deficits observed in Mib2 mice during behavioral experiments and electrophysiological recordings were due to the dysregulation of GluN2B caused by the absence of Mib2. However, we found similar levels of pGluN2B and GluN2B in Mib2 KO and WT mouse hippocampi. In our study, we used whole hippocampal lysates. If Mib2 ubiquitinates GluN2B subunits only at the postsynaptic, but not at the extrasynaptic site, the misregulation of postsynaptic GluN2B ubiquitination could be obscured by the non-affected extrasynaptic GluN2B population. In another report [16], Mib2 was shown to regulate the Notch signaling pathway by ubiquitinating the ligand protein Delta. Although Notch signaling is mainly known to regulate development [34], recent studies showed that it is also activated by neuronal activity and regulates synaptic plasticity [12]. Because Notch signaling in neuronal cells regulates activity-dependent synaptic plasticity [1135], the impaired memory and synaptic plasticity in Mib2 KO mice might be due to a decreased Notch signaling. More importantly, our previous report showed that conditional knock-out (cKO) of Mib1, a Mib2 paralogue, in mature neurons of the mouse forebrain results in impaired Notch signaling [18]. To test if Mib2 is also involved in Notch signaling in the adult mouse brain, we measured the level of cleaved Notch1 in the hippocampi of Mib2 KO and WT mice. Unlike the forebrain of Mib1 cKO mice [18], Mib2 KO and WT mouse hippocampi had a comparable level of cleaved Notch1. However, when cleaved Notch1 levels were measured 1 hour after mild foot shock, Mib2 KO mouse hippocampi contained a lower level of cleaved Notch1 than WT hippocampi. Mib2 is highly expressed in adult brain tissues, but shows a lower expression level in embryonic brain tissues; however, Mib1 is constantly expressed at a high level [16]. Therefore, our data indicate that Mib1 is involved in basal Notch signaling in the adult brain, while Mib2 is involved in neural activity-dependent Notch signaling. However, since Mib2 KO mice are conventional KO mice line, the possibility that normal levels of basal Notch signaling was maintained by a compensatory mechanism in Mib2 mice still remains.
The lower level of cleaved Notch1 after fear conditioning and the impaired synaptic plasticity in Mib2 KO mice implicates that Mib2 might mediate Notch signaling during activity-dependent synaptic plasticity in the hippocampus. Our study suggests that the absence of Mib2 leads to decreased hippocampal Notch signaling during learning, and thus causes impaired hippocampus-dependent learning, memory formation, and synaptic plasticity in Mib2 KO mice.
 XML Download
XML Download