

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
The oral cavity is continuously exposed to elements of both the internal and external environments, and is populated by beneficial commensal microorganisms as well as many pathogens. Human gingival epithelial cells are the first line of defense against exogenous stimuli such as bacterial toxins, allergens, thermal change, and osmotic molecules. The gingival epithelial barrier is adjacent to various cells of the immune system such as T cells, dendritic cells, neutrophils, macrophages, and mast cells, ready to initiate an immune response [1]; furthermore, the oral epithelium secretes cytokines in response to harmful microorganisms [2,3,4]. When microorganisms invade, the innate immune system of the host recognizes pathogens through pattern-recognition receptors [5] such as toll-like receptors (TLRs) on epithelial cells, neutrophils, monocytes, dendritic cells, and fibroblasts [6,7,8]. It is important to maintain an appropriate balance between commensal bacterial colonization and local immune responses in the oral cavity.
The main etiological feature of periodontal disease is the presence of anaerobic gram-negative bacteria in the subgingival area such as Actinobacillus actinomycetemcomitans, Porphyromonas gingivalis, Prevotella intermedia, and Tannerella forsythensis [9]. Lipopolysaccharide (LPS) produced by gram-negative bacteria appears to contribute to the progression and severity of periodontal disease [10]. Furthermore, gram-positive as well as gram-negative bacteria have been shown to induce oral inflammation [11,12]. Lipoteichoic acid, found in gram-positive bacteria, contributes to E. faecalis-induced periodontitis [11], and high levels of the gram-positive bacteria E. nodatum and S. exigua have been associated with clinical indicators of periodontal disease [12].
In gram-positive bacteria, peptidoglycan (PGN) makes up as much as 90% of the bacterial cell wall, which is the outermost structure recognized by TLR2 [5,13]. Like LPS, PGN is an important bacterial component with respect to periodontal disease. TLRs, which are expressed on oral epithelial cells and several immune cells, including macrophages, dendritic cells, and B cells, play a crucial role in the detection of microbial infection in mammals and insects [3,14]. To date, ten TLR family members have been identified in the human genome, and several TLRs activate the early innate immune response [13]. TLR2 signaling is mediated by adaptor proteins such as myeloid differentiation 88 (MyD88) and Toll/interleukin-1 (IL-1)-receptor (TIR)-domain and used in the signaling cascade [13,15]. TLR2 is also involved in the secretion of pro-inflammatory chemokines and other cytokines. PGN-induced TLR2 activation induces interleukin-6 (IL-6) and interleukin-8 (IL-8) [16,17,18]; however, the exact intracellular mechanism of TLR2 activation has yet to be elucidated.
Intracellular calcium ([Ca2+]i) plays an important role in cellular functions that regulate gene expression, growth, differentiation, apoptosis, muscle contraction, memory, and learning. Moreover, Ca2+ signaling controls target gene activation and induces downstream immune responses and inflammation [19,20,21,22]. Research has shown that PGN increases [Ca2+]i through recruitment of phosphatidylinositide 3-kinase (PI3K) and phospholipase Cγ2 (PLCγ2) to affect the release of Ca2+ from intracellular stores in macrophages and dendritic cells [23,24]. Moreover, activation of Ca2+ signaling induces the secretion of IL-8, indicating that the inflammatory response is directly affected by a wide range of activation pathways [23,25].
When gingival epithelium is exposed to pathogens, it is unknown whether bacterial PGN triggers Ca2+ release, and if so, from which Ca2+ store. In the present study, we investigate the direct effect of PGN on Ca2+ signaling and IL-8 expression in human gingival epithelial cells.
Go to : 
METHODS
Reagents
Keratinocyte basal medium-2 (KBM-2) was purchased from Lonza (Walkersville, MD). Collagenase A and dispase II were obtained from Roche (Mannhein, Germany) and U73122, U73344, PGN, and LPS were the products of Sigma (St. Louis, MO). Thapsigargin (Tg) was obtained from Alexis Biochemical (San Diego, CA). Fetal bovine serum (FBS), fura-2-acetoxymethyl ester (fura-2,AM), and Pluronic F-127 were purchased from Invitrogen (Carlsbad, CA). All other chemicals were purchased from Sigma.
Cell culture
All experimental protocols were reviewed and approved by the Research Ethics Committee of Yonsei University College of Dentistry and Dental Hospital. Informed consent was obtained from all patients according to the requirements of the Institutional Review Board. Human gingival epithelial cells were cultured from the explant tissues of healthy donors who underwent third molar extraction at Yonsei University Dental Hospital. The gingival epithelium was gently separated from connective tissues by treatment with collagenase A and dispase II, and then cultured in KBM-2 containing 10% FBS and 1% antibiotics in a 5% carbon dioxide (CO2) incubator at 37℃. All experiments were carried out with 3 to 4 passages of the human gingival cells.
Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
Total RNA was prepared using Trizol reagent (Invitrogen) according to the manufacturer's instructions. RNA concentrations were determined by measuring the amount of UV absorption at 260 and 280 nm. Total isolated RNA was amplified according to the manufacturer's protocol using AccuPower® RT PreMix (Bioneer, Seoul, Korea). Samples subjected to amplification without reverse transcriptase served to verify the absence of genomic DNA. The cDNA was amplified by PCR with HiPi™ Thermostable DNA polymerase (Elpis, Seoul, Korea). Primers used were as follows: for GAPDH (307 bp), 5'-CGG AGT CAA CGG ATT TGG TCG TAT-3'(forward), 5'-AGC CTT CTC CAT GGT GGT GAA GAC-3'(reverse); for IL-8 (292 bp), 5'ATG ACT TCC AAG CTG GCC GTG GCT-3'(forward), 5'-TCT CAG CCC TCT TCA AAA ACT TCT-3'(reverse). The PCR program began with a 5-min denaturation at 95℃, followed by 35 cycles of 95℃/1 min, 58℃/1 min, 72℃/1 min. PCR samples were electrophoresed on 1.2% agarose gels in TAE (tris-acetate-ethylenediaminetetraacetic acid electrophoresis) buffer. The quantity of PCR-generated DNA fragments was estimated relative to DNA ladder standards.
Measurement of [Ca2+]i
Human gingival epithelial cells were cultured on collagen-coated cover-slipped slides. Cells were loaded with 5 µM fura-2-AM in the presence of 0.05% Pluronic F-127 for 30 min in physiological salt solution (PSS). PSS composition was as follows: 140 mM sodium chloride, 5 mM potassium chloride, 1 mM magnesium chloride, 10 mM 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid, 1 mM calcium chloride, and 10 mM glucose, titrated to pH 7.4 with sodium hydroxide and with an osmolarity of 310 mOsm. Changes in [Ca2+]i were measured by means of fura-2-AM fluorescence with excitation wavelengths of 340 and 380 nm and an emission wavelength of 510 nm. The emitted fluorescence was monitored with a charge-coupled device camera and analyzed with a MetaFluor system (Molecular Devices, PA). Fluorescence images were obtained at 2-sec intervals. Background fluorescence was subtracted from the raw signals of background at each excitation wavelength before calculating the fluorescence ratio as F340/F380 as described previously [26].
Statistical analyses
Data from at least three independent experiments were expressed as mean±standard error (SE). Statistically significant differences between groups were determined using the paired student's t-test.
Go to :
RESULTS
PGN induces Ca2+ signaling in a concentration-dependent manner and increases IL-8 mRNA expression in human gingival epithelial cells
To investigate Ca2+ signaling evoked by PGN, [Ca2+]i was measured at PGN concentrations ranging from 1 to 125 µg/ml. PGN induced increases in [Ca2+]i in a concentration-dependent manner (Fig. 1A). The amplitude of the peak was affected by PGN concentration from the basal line across the test concentration range of 1 to 125 µg PGN/ml, with a half maximal effective concentration (EC50) of 22.45 µg PGN/ml (Fig. 1B). LPS and PGN treatment induced tyrosine phosphorylation of phospholipase Cγ-2 in bone marrow-derived macrophages and dendritic cells [23]. Cells were treated with LPS and PGN for 24 hr and 1 hr, respectively, and IL-8 mRNA expression levels were measured. Compared with control cells, PCR analysis revealed 1.6- and 1.8-fold higher IL-8 mRNA expression in LPS- and PGN-treated cells, respectively (Fig. 1C).
 | Fig. 1PGN-induced Ca2+ responses in human gingival epithelial cells. (A) Changes in [Ca2+]i induced by concentrations of PGN ranging from 1 to 125 µg/ml (n=3). (B) Analysis of concentration-dependent changes in the peak of fluorescence based on the F340/F380 ratio. (C) Human gingival epithelial cells were stimulated with LPS (100 µg/ml) and PGN (50 µg/ml). Total mRNA was extracted from the cells, and IL-8 mRNA expression was measured (n=3). Data are presented as mean±SE. *p<0.05; **p<0.01 versus control.
|
PGN induces Ca2+ release from the endoplasmic reticulum (ER), and PGN-induced IL-8 expression is regulated by the phospholipase C/ inositol 1,4,5-trisphosphate (PLC/IP3) pathway in human gingival epithelial cells
Ca2+ signaling is mediated by several Ca2+ sources. To determine the source(s) of PGN-induced [Ca2+]i release, 50 µg/ml PGN was added to the Ca2+-free extracellular medium and chelated with 10 mM ethylene glycol tetraacetic acid. PGN induced [Ca2+]i increases in the absence of extracellular Ca2+ (Fig. 2A), suggesting that the increased [Ca2+]i originates from intracellular Ca2+ stores, most probably from ER. The ER Ca2+ stores can be depleted by Tg, a specific inhibitor of the sarco/endoplasmic reticulum Ca2+ ATPase (SERCA) pump. Accordingly, cells were pretreated with PGN for 7 min and the Ca2+ store was depleted with the addition of 1 µM Tg in a nominally Ca2+-free medium. After depletion of the Tg-sensitive Ca2+ store, PGN failed to evoke a Ca2+ signal (Fig. 2B, left). Furthermore, after depletion of Ca2+ stores by PGN, Tg treatment failed to evoke a Ca2+ signal (Fig. 2B, right). In addition, to determine the role of the PLC/IP3 pathway in PGN-induced [Ca2+]i increases, cells were pre-treated with 10 µM U73122, a specific blocker of PLC, or its inactive analog, 10 µM U73343 for 5 min. U73122 effectively blocked PGN-induced [Ca2+]i increases (Fig. 2C, left) whereas U73343 produced no effect (Fig. 2C, right), suggesting that PGN increases [Ca2+]i by Ca2+ release from the ER through the PLC pathway. Cells also were treated with the IP3 receptor (IP3R) blocker, 2-aminoethoxydiphenyl borate (2APB, 75 µM), to determine whether PGN-induced [Ca2+]i increase is mediated by IP3R. As shown in Figure 2D, treatment with 2APB during PGN stimulation inhibited [Ca2+]i increase, suggesting that PGN increases [Ca2+]i by promoting Ca2+ release from the ER through the PLC/IP3 pathway. Furthermore, we examined the effects of PGN on IL-8 mRNA expression following pre-treatment with U73122 or 1,2-bis (2-aminophenoxy) ethane-N,N,N',N'-tetraacetic acid tetrakis-AM (BAPTA-AM). PGN-induced IL-8 mRNA expression was decreased following U73122 or BAPTA-AM treatment (0.6-fold decrease) (Fig. 2E). These results suggest that PGN-induced IL-8 expression is regulated by the PLC/IP3 pathway.
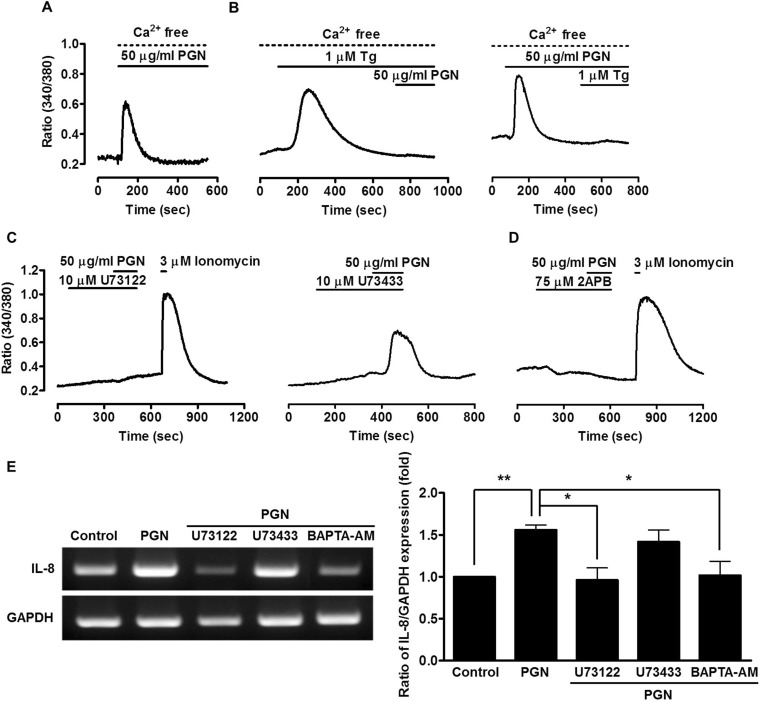 | Fig. 2Characterization of PGN-induced Ca2+ signaling. (A) Effect of removal of extracellular Ca2+ on PGN-induced increase in [Ca2+]i (n=3). (B) Effect of Tg (1 µM) on PGN-induced [Ca2+]i increases in the absence of extracellular Ca2+ (n=3). (C) Effect of U73122 (10 µM) and U73343 (10 µM) on PGN-induced increase in [Ca2+]i (n=3). Ionomycin was used as a positive control. (D) Effect of 2APB (75 µM) on PGN-induced increase in [Ca2+]i (n=3). (E) Oral gingival epithelial cells were treated with U73122, U73343, or BAPTA-AM for 20 min and stimulated with PGN for 1 h and then total RNA was extracted from the cells. Data are presented as mean±SE. *p<0.05; **p<0.01 versus control.
|
Ca2+ signaling significantly increases IL-8 mRNA expression
The main source of PGN-induced Ca2+ release is the ER. Intracellular Ca2+, especially store-operated Ca2+ movements, mediates long- and short-term inflammation and cytokine release [27,28]. To assess whether levels of pro-inflammatory cytokines such as IL-8 increase with increasing Ca2+, gingival cells were treated with 1 µM Tg for 1 h and IL-8 mRNA expression levels were measured. Figure 3, shows that IL-8 mRNA expression in cells treated with Tg was 1.4-fold higher than in controls. Additionally, co-treatment with Tg and PGN increased IL-8 mRNA expression (non-additively, however). These results indicate that PGN induces the increase of IL-8 mRNA expression mediated by intracellular Ca2+ levels through the Tg-sensitive ER store in human gingival epithelial cells.
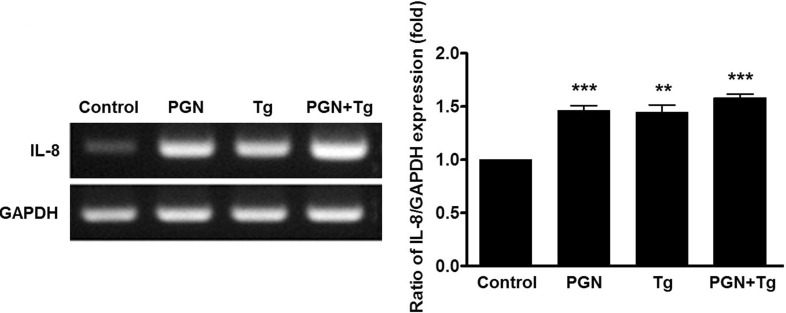 | Fig. 3Effect of Ca2+ signaling on IL-8 mRNA expression. Oral epithelial cells were stimulated with Tg and PGN for 1 h and then total RNA was extracted from the cells. Data are presented as mean±SE of values from four independent experiments. Data are represented as mean±SE. **p<0.05; ***p<0.01 versus control.
|
Go to :
DISCUSSION
Periodontal disease results from interactions between pre-disposed host tissue and bacterial plaque. Human gingival epithelial cells, as a susceptible host, represent the first line of defense against exogenous stimuli. The human mouth is continuously occupied by a complex array of microorganisms, including Gram bacteria, fungi, and viruses [29]. Gram-negative bacteria in particular are considered one of the major contributors to periodontal diseases. Studies have also revealed that gram-positive bacteria play a significant role in periodontal disease [3,12]. It is unknown whether cytokine secretion in response to gram-positive bacteria affects the inflammation of oral gingival epithelial cells.
PGN fragments have been shown to be a natural ligand for the nucleotide-binding oligomerization domain-containing protein 2 (NOD2) receptor in immune cells [30]. NOD2-mediated activation of dendritic cells with polymeric PGN is dependent on TLR2 co-stimulation [31]. It is still unclear whether PGN interacts with TLR2; however, stimulation of mouse keratinocytes with PGN from Staphylococcus aureus SA113 resulted in co-localized TLR2 and NOD2 receptors and induced host immune responses [32]. Still, the role of PGN, its potent receptors, and their effects in gingival epithelial cells are unclear.
In the present study, we demonstrated for the first time that PGN increases [Ca2+]i in human gingival epithelial cells in a concentration-dependent manner via intracellular Ca2+ release (Fig. 1A and B). The peptidoglycan caused many pathological changes demonstrated by disruption of alveoli walls, edema, and degeneration of cells lining the bronchioles. Peptidoglycan was used 37×104 µg/ml to compare the pathological effect of 1×109 cfu/ml bacterial suspension for an injection to the mouse [33]. We demonstrated that peptidoglycan increases [Ca2+]i in human gingival epithelial cells and those concentration ranges of peptidoglycan induced an increased proinflammatory cytokine IL-8 mRNA expression. Based on the Fig. 1B, those concentration ranges can be considered with physiological and pathological condition. PGN induced Ca2+ release from Tg-sensitive ER stores (Fig. 2). These results are consistent with previous studies that have shown that PGN induced tyrosine phosphorylation of PLCγ2, leading to intracellular free Ca2+ mobilization in macrophages and dendritic cells [23]. We also showed that PGN stimulated the expression of pro-inflammatory cytokine IL-8. IL-8 is a potent chemo-attractant produced by macrophage and other cell types such as epithelial cells, fibroblasts, and endothelial cells [34]. We found that PGN-induced expression of IL-8 mRNA is inhibited PLC/IP3 pathway antagonists (Fig. 2E). Previous research has shown that, in PLCγ2-knockdown cells, PGN-induced phosphorylation of nuclear factor of kappa light polypeptide gene enhancer in B-cells inhibitor, alpha (IκB-α) and p38 activation were reduced [23]. Pro-inflammatory signals such as nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) signaling plays a central role in inflammation [21,35]. PGN-induced activation of the TLR2/MyD88 main pathway activates NF-κB signaling [13,15]; PGN-induced increases in [Ca2+]i can also activate NF-κB signaling [23,24]. Furthermore, previous studies have shown that NF-κB activation is regulated by intracellular Ca2+ concentrations [21]. To investigate whether intracellular Ca2+ can regulate cytokine expression, we measured IL-8 mRNA expression after Tg stimulation (which has been shown to elevate intracellular Ca2+ concentrations in various cell types), and found that Tg induced increases in IL-8 mRNA expression (Fig. 3A). Previous research has reported that store-operated Ca2+ channels (SOCs) play an essential role in the short- and long-term regulation of the inflammatory process [27]. IP3R releases Ca2+ from the ER; depletion of Ca2+ in the ER stores also activates SOCs. The transcriptional regulation of IL-8 expression in epithelial cells is complex and involves many other factors such as nuclear factor NF-κB, NF-IL 6, and activator protein (AP)-1 [36]. We have confirmed direct effects of peptidoglycan on IL-8 mRNA expression. Moreover peptidoglycan-induced IL-8 mRNA expression was decreased following U73122 or BAPTA-AM treatment. Thus IL-8 mRNA expression level is sufficient to explain the direct effect of peptidoglycan as recommended with/without several modulators of calcium signals. We suggest a correlation between intracellular Ca2+ and cytokine expression in human gingival epithelial cells.
In the present study, we show that PGN induced increases in [Ca2+]i through the PLC/IP3 pathway and consequently increased IL-8 mRNA expression. Furthermore, IL-8 mRNA expression was dependent on intracellular Ca2+ levels in human gingival epithelial cells. Therefore, the development and use of inhibitors of intracellular Ca2+ might provide new modalities for the treatment of periodontal disease.
Go to :
 XML Download
XML Download