

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Microglia, the resident immune cells in the CNS, constitute 5~20% of the total cells in the mammalian brain. Microglia can be activated in response to even minor pathological changes and transform function as well as morphology [12]. These activated microglia phagocytose dead cells and cellular debris [3]. Studying microglia in vitro and ex vivo, however, has a limitation in elucidating the in vivo microglia characteristics because the slicing and culture procedure itself can activate microglia [34]. Whereas, in vivo two-photon imaging studies allowed us to directly monitor the 'resting' microglia in an intact brain, and revealed that resting microglia are also highly motile; their processes continuously extend and retract to survey the microenvironment for surveillance, and once they encounter tissue damage, microglia rapidly respond by extending their processes to the injury site in the CNS [56].
Toll-like receptor 2 (TLR2) is an innate immune receptors recognizing bacterial lipoteichoic acid and lipoprotein, and triggering inflammatory responses in innate immune cells [7]. TLR2 is widely expressed in the central nervous system (CNS), particularly in microglia [8]. Previously, we have reported that TLR2 plays a critical role in microglia activation in various neurological diseases [9]. TLR2 is required for the nerve injury-induced spinal cord microglia activation and subsequent neuropathic pain development [10]. In addition, TLR2 contribute to microglia activation and hippocampal neurodegeneration in kainic acid-induced epilepsy model [11], and also in traumatic brain injury [12]. Therefore, it is likely that TLR2 is utilized by microglia to detect brain tissue damage. However, its role in immediate microglial reaction to brain tissue injury in vivo does not have been resolved. In the present study, we used in vivo two-photon microscopy and Cx3cr1GFP/+ knock-in/TLR2knock-out (KO) mice to investigate whether TLR2 plays a role in the immediate early microglial motility responding to a two-photon laser-induced local injury in the intact cortex.
METHODS
Animal preparation
All procedures were approved by the Institutional Animal Care and Use Committees of Kyung Hee University (KHUASP(SE)-14-016) and Seoul National University (SNU-140408-16). Cx3cr1GFP/+ knock-in mice were obtained from the Jackson Laboratory (Bar Harbor, USA), and TLR2 KO mice were obtained from Dr. Akira (Osaka University, Osaka, Japan).
Adult (2~4 months old) Cx3cr1GFP/+ knock-in mice and Cx3cr1GFP/+ knock-in/TLR2-KO mice were used to visualize microglia. Mice were deeply anesthetized with an intraperitoneal injection of urethane (1.64 g/kg). The animal skull was exposed above the somatosensory cortex (1 mm posterior to bregma and 2 mm lateral) and cleaned. A small circular craniotomy (~2 mm in diameter) was then carefully performed using a high-speed drill, which was covered with a thin glass cover slip [1314]. For thinned-skull cranial window, the skull was thinned with high-speed drill and scraped with a microsurgical blade to a thickness of ~20 µm [15].
In vivo two-photon microscopy imaging
Imaging was performed with a two-photon microscope (Zeiss LSM 7 MP, Carl Zeiss). A Ti:sapphire laser (Chameleon, Coherent) was tuned to the excitation wavelength for GFP (900~950 nm) [13]. Subsequent image stacks (512×512 pixels, 0.4 µm/pixel, 26 sections) were recorded every 5 min for 1~2 hour. The imaging depth was 50~150 µm from the pial surface.
Two-photon laser ablation
Laser ablation was induced by focusing a two-photon laser beam (800 nm, 70 mW) in the somatosensory cortex (target: microglia soma, spot size: 15×15 µm) to create a small injury site as indicated by an autofluorescent sphere [6].
Data analysis
Image processing was done using ImageJ (http://rsbweb.nih.gov/ij). The length of processes was evaluated from maximum-intensity projections and the motility was calculated as absolute Δlength in comparison between two successive imaging sessions.
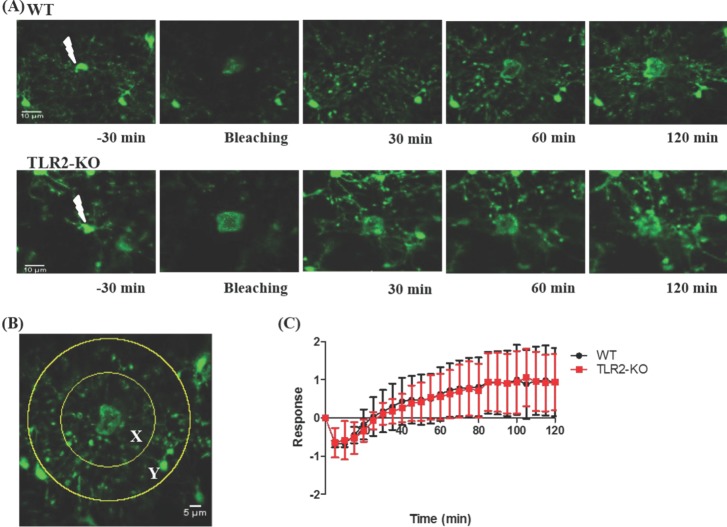
In laser ablation experiments, we quantified the degree of the invasion of microglial processes to the injured site as described previously [6]. Briefly, the number of microglial processes entering from the outer region Y (70 µm in radius) into the inner region X (35 µm in radius) surrounding the lesion site was measured before and after the laser ablation. We counted the pixels containing GFP signal in the region X over time [Rx(t)], from which Rx(0), the GFP pixels in X immediately after the laser ablation, was subtracted. This subtracted value was then divided by Ry(0), the GFP pixels in the region Y immediately after the ablation. The microglial response, R(t), is thus provided by R(t)=(Rx(t)-Rx(0))/Ry(0).
All data are presented as mean±SEM. Statistical analysis and graphic works were done with Prism 5.0 (Graph Pad Software, USA). Unpaired t test or two-way repeated measures analysis of variance (ANOVA) followed by Dunnett's multiple comparison test were used. In all cases, p<0.05 was considered significant.
RESULTS
The motility of microglial processes in WT and TLR2-KO mice
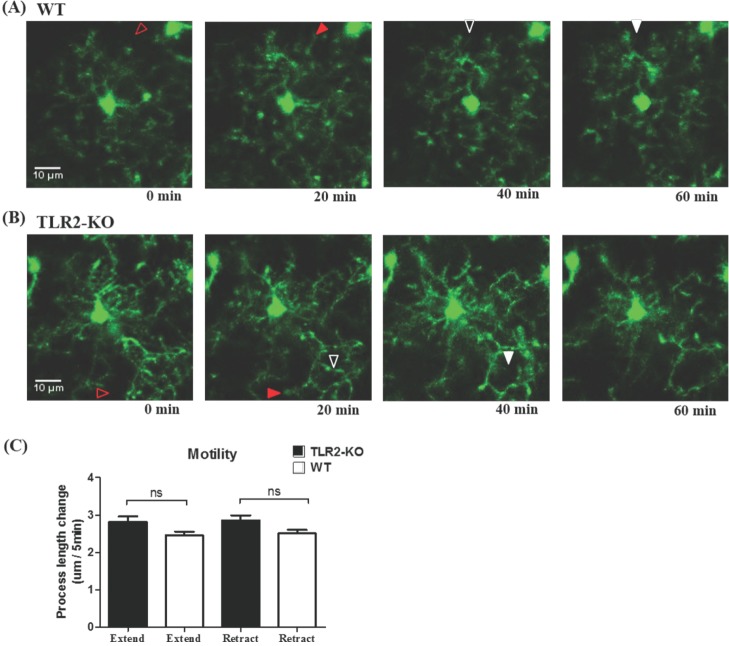
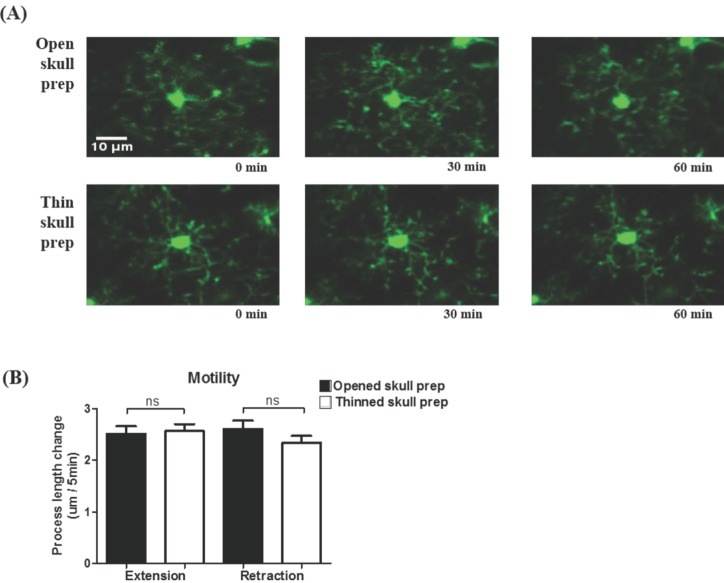
By using in vivo two-photon microscopy imaging through a cranial window, we first observed the resting state dynamics of microglial processes for an hour in the intact somatosensory cortex of TLR2-KO or WT mice. As shown in Fig. 1A and 1B, the resting microglia in the both groups of mice had a small rod-shaped soma and ramified processes. Their processes continually extended and retracted, but microglial soma remained fixed (Fig. 1A and 1B). To quantify the motility (i.e. extension and retraction) of resting microglial processes, we measured the length changes of individual processes every five minutes. Fig. 1c shows that there was no significant difference in the motility of microglial processes between the TLR2-KO and WT mice (p>0.05). Imaging through a thinned-skull cranial window showed a similar shape and motility of microglia in WT mice to that obtained through an open-skull window (Fig. 2), indicating that our craniotomy procedure does not alter a physiological condition of microglia.
Microglia rapidly respond to local brain injury in TLR2-KO and WT
By using a two-photon laser ablation method, we next examined an immediate response of microglia to a local brain injury in TLR2-KO and WT mice. As shown in Fig. 3A, this local injury was developed after a focal two-photon laser stimulus was applied to a microglia soma, and then nearby microglia rapidly sent out their processes to the lesion site in both groups of mice. More specifically, within a few minutes after a laser ablation, the microglial processes close to the damaged site appeared slightly enlarged and bulbous. After the next few minutes, microglial processes reached to the damaged site (Supplementary Videos 1 and 2). To quantitatively compare the microglial response to laser-induced injury in TLR2-KO and WT mice, we counted the number of pixels entering from the outer area Y into the inner area X with time (Fig. 3B and see also Methods). Fig. 3C shows that there was no significant difference in the microglial response to the injury between the TLR2-KO and WT mice (p>0.05).
DISCUSSION
In the present study, we first investigated whether the 'resting' state microglia behave differently between the TLR2-KO and WT mice. Irrespective of the presence of TLR2 signaling, their processes continually extended and retracted, in consistent with a previous report [5], with no significant difference in the motility between the two groups of mice (Fig. 1). These results indicate that the microglial behavior in the resting state is not dependent on TLR2 signaling. Although an open-skull preparation for long-term in vivo two-photon imaging is reported to gradually cause glial activation [16], our experimental conditions using acute and short-term two-photon imaging of microglia that located 50~150 µm from the pial surface showed no difference in the morphology and motility of resting microglia between an thinned-skull preparation and an open-skull window (Fig. 2).
Our previous studies have reported that TLR2 has a critical role in microglial activation and pathogenesis in the CNS [101112]. Other studies also demonstrated that TLR2-mediated signaling promotes the production of multiple inflammatory mediators such as interleukin (IL)-1, IL-6, tumor necrosis factor (TNF)-α and nitric oxide (NO) [171819]. In addition, axonal injury in the mouse brain induced early cytokine/chemokine expressions (e.g. TNF-α, CXCL10), T cell infiltration and microglial expansion, all of which were significantly reduced in TLR2-KO mice [20]. These results indicate that microglia activated through TLR2 signaling induces neuroinflammation and neurite degeneration. In contrast, a recent ex vivo study [21] showed that a TLR2 agonist, Pam2CSK4, increased the microglial response to laser-induced spinal cord injury, reduced proximal axonal dieback and secondary axonal degeneration, and induced an alternative microglial activation profile, suggesting that this TLR2-mediated microglial activation might be protective. However, other TLR2 agonists had little effect on such microglial responses and axon degeneration, and the contrary effect of TLR2-KO seemed to be very slight [21]. Thus, the role of TLR2-mediated microglial activation in the pathological states is still controversial. Furthermore, it remains to be unclear whether TLR2 has a role in immediate microglial reaction to a local brain injury in vivo.
In the second set of the experiments, we took advantage of the focal properties of the two-photon microscopy laser [6] to identify how microglia immediately react to local brain injury in TLR2-KO and WT mice. Unexpectedly, we found that TLR2-KO mice exhibited a similar microglial response to the injury when compared to that of WT mice (Fig. 3). These results also indicate that the immediate microglial reaction to a local brain injury is not dependent on TLR2 signaling.
In conclusion, this study demonstrates that the microglial process of TLR2-KO mice showed similar level of motility to that of WT mice in the 'resting' state. Following a two-photon laser ablation-induced focal injury, the microglial response motility to the injury is also similar between WT and TLR2-KO mice. Therefore, our findings suggest that TLR2 signaling does not have an essential role in the resting state microglial behavior as well as in the immediate early microglial response to a brain tissue injury in vivo.

 XML Download
XML Download