

 PDF
PDF ePub
ePub Citation
Citation Print
Print


ABBREVIATIONS
CDK
cyclin-dependent kinase
ERK1/2
extracellular signal-regulated kinase 1/2
MAPK
mitogen-activated protein kinase
PCNA
proliferating cell nuclear antigen
PDGF
platelet-derived growth factor
PI3K
phosphatidylinositol-3-kinase
Akt/PKB
protein kinase B
PLCγ1
phospholipase Cγ1
Rb
retinoblastoma
STAT3
signal transducer and activator of transcription 3
VSMC
vascular smooth muscle cell
INTRODUCTION
Abnormal proliferation of vascular smooth muscle cells (VSMCs) is a key feature of vascular disorders, including restenosis and neointimal hyperplasia [1]. VSMC proliferation and migration are induced by several cytokines and growth factors. One of the major stimulants of the mitogenesis of VSMCs is platelet-derived growth factor (PDGF) [2]. PDGF is secreted from damaged blood vessel walls and stimulates cell proliferation and migration through signal transduction pathways [3]. PDGF, which is produced by activated macrophages, VSMCs, and endothelial cells, exists in three isoforms (AA, AB, and BB). PDGF-BB is a considerably more potent proliferative stimulus for VSMCs than PDGF-AA [4]. PDGF-BB, a growth factor secreted by injured endothelial cells, VSMCs, platelets, and macrophages, promotes the proliferation of fibroblasts, glial cells, and VSMCs [5].
The binding of PDGF-BB to PDGF receptor leads to dimerization and autophosphorylation of tyrosine residues, resulting in the downstream activation of phosphatidylinositol-3-kinase (PI3K), phospholipase C (PLC)γ1, and/or mitogen-activated protein kinase (MAPK) pathways [67]. Signal transducer and activator of transcription 3 (STAT3) is also thought to participate in PDGF-induced cell proliferation [89]. Activation of these mitogenic signals triggers cell cycle progression [1011].
Cellular proliferation is regulated by the cell cycle, which consists of four distinct sequential phases: G0/G1, S, G2, and M. This tightly regulated temporal order is controlled by the sequential activation of serine/threonine protein kinases, known as cyclin-dependent kinases (CDKs), which phosphorylate the retinoblastoma protein (pRb). The cell cycle transition is controlled by the action of CDKs and their activating subunits, cyclins [1213]. After vascular injury, VSMCs are stimulated to divide in response to mitogens, and they exit G1 phase and enter S phase. Cyclin D1/CDK4 and cyclin E/CDK2 complexes perform important roles in promoting the transition from G0/G1 to S phase and are required for cell cycle progression through this period [14].
Murrayafoline A, a carbazole alkaloid isolated from Glycosmis stenocarpa Guillamin (Rutaceae), has been reported to possess various pharmacological activities, including antifungal and anticancer properties [1516]. However, limited studies have been published on the effects of murrayafoline A on VSMCs. The present study was designed to investigate the effects of murrayafoline A on PDGF-BB-induced VSMC proliferation and the cell cycle, and to determine the underlying molecular mechanism(s) responsible for these effects.
Go to : 
METHODS
Test compound and other materials
Murrayafoline A [Brown oil, C14H13NO, Rf: 0.25 (hexane/EtOAc, 10: 0.5), EI-MS m/z: 211 (100%) 196 (M-CH3)+, 167, 139, 115, 101, 77] was obtained as previously described [17]. The structure of murrayafoline A was established by 1H- and 13C-NMR analysis. The purity of murrayafoline A was estimated to be higher than 97% by both HPLC and spectroscopic analysis. All cell culture materials were purchased from Invitrogen (Carlsbad, CA, USA). Anti-phospho-ERK1/2, anti-phospho-PLCγ1, anti-phospho-PDGF-R β (Tyr751), anti-phospho-STAT3 (Tyr705), anti-ERK1/2, anti-Akt, anti-PLCγ1, and anti-PDGF-Rβ antibodies were purchased from Cell Signaling Technology, Inc. (Beverly, MA, USA). Anti-phospho-Akt antibodies were purchased from Millipore Corp. (Billerica, MA, USA). Anti-phospho-pRb, anti-CDK2, anti-CDK4, anti-phospho PCNA, anti-cyclin D1, anti-cyclin E, anti-Akt, and anti-β-actin antibodies were purchased from Abfrontier (Geumcheon, Seoul, Korea). PDGF-BB was obtained from Upstate Biotechnology (Lake Placid, NY, USA). All other chemicals used were of analytical grade.
Cell culture
Rat aortic VSMCs were isolated by enzymatic dispersion as described previously [18]. Cells were cultured in Dulbecco's Modified Eagle Medium (DMEM) supplemented with 10% fetal bovine serum (FBS), 100 IU/ml penicillin, 100 µg/ml streptomycin, 8 mM HEPES, and 2 mM L-glutamine at 37℃ in a humidified atmosphere of 95% air and 5% CO2. The purity of the VSMC culture was confirmed by immunocytochemical localization of β-smooth muscle actin. The VSMCs used in these experiments were of passages 4-8.
Cell proliferation assay
VSMC proliferation was measured by both direct counting and a non-radioactive colorimetric WST-1 assay (premix WST-1; Takara Bio Inc., Otsu, Japan). For direct cell counting, VSMCs were seeded into 12-well culture plates at 4×104 cells/ml, and then cultured in DMEM containing 10% FBS at 37℃ for 24 h. After reaching ~70% confluence, the cells were incubated with serum-free medium for 24 h, treated with various concentrations of murrayafoline A for another 24 h in new, fresh serum-free medium, and stimulated with PDGF-BB (50 ng/ml). Murrayafoline A was dissolved in dimethyl sulfoxide (DMSO); the final concentration of DMSO in the medium did not exceed 0.1%. After 24 h, the cells were trypsinized with trypsin-EDTA and counted using a hemocytometer. For the non-radioactive colorimetric WST-1 assay, all procedures were performed according to the manufacturer's protocol, and the results are expressed as a percentage of the control.
Cell viability assay
VSMCs were seeded in 96-well culture plates at 3×104 cells/ml and then cultured in DMEM containing 10% FBS at 37℃ for 24 h. When the cells reached ~70% confluence, they were incubated with serum-free medium for another 24 h and then exposed to 5 µM murrayafoline A or 100 µg/ml digitonin as a cytotoxic control for the given time [19]. After 2 h of incubation with the WST-1 reagent, the absorbance was measured at 450 nm using a microplate reader (Packard Instrument Co., Downers Grove, IL, USA).
DNA synthesis assay
DNA synthesis was determined using a [3H]-thymidine incorporation assay. The assay conditions were as described for the cell proliferation assay. Under stimulatory conditions, PDGF-BB (50 ng/ml) added to serum-free medium, [3H]-thymidine (2 µCi/ml) was added for 4 h before harvesting. The reaction was terminated by aspirating the medium and subjecting the cultures to sequential washes on ice with PBS containing 10% trichloroacetic acid and ethanol/ether (1:1, v/v). Acid-insoluble [3H]-thymidine was extracted into 250 µl of 0.5 M NaOH/well, and this solution was mixed with 3 ml of scintillation cocktail (Ultimagold; Packard Bioscience, Meriden, CT, USA) and quantified using a liquid scintillation counter (LS3801; Beckman, Düsseldorf, Germany).
Cell cycle progression analysis
Cell cycle progression assay conditions were as described for the cell proliferation assay. After stimulation with PDGF-BB (50 ng/ml) for 24 h, cells were trypsinized and centrifuged (1,500×g, 7 min). The pellets were resuspended in 1 ml of 1×PBS, washed twice, and fixed with 70% ethanol for 48 h. The fixed cells were briefly vortexed and centrifuged (15,000×g, 5 min). The ethanol was discarded and the pellets were stained with 500 µl of propidium iodide (PI) solution (50 µg/ml PI in sample buffer containing 100 µg/ml RNase A). Before analysis by flow cytometry, each sample was incubated at room temperature for 1 h. The PI DNA complex in each cell nucleus was measured with a FACSCalibur (Becton, Dickinson and Co., Franklin Lakes, NJ, USA). The nuclear DNA content was reflected by the fluorescence intensity of the incorporated PI. The numbers of cells in G0/G1, S, and G2/M phase were determined using ModFit LT software.
Immunoblotting
VSMCs were stimulated with 50 ng/ml PDGF-BB for the phosphorylation of PDGF-Rβ (3 min), extracellular signal-regulated kinase 1/2 (ERK1/2, 5 min), PLCγ1 (5 min), STAT3 (10 min), and Akt (15 min). For the detection of cyclin D1, cyclin E, CDK2, CDK4, and PCNA expression, and pRb phosphorylation, cells were stimulated with 50 ng/ml PDGF-BB for 24 h. Immunoblotting was performed as previously described [20].
Statistical analyses
All data are expressed as means±SEM. A one-way ANOVA was used for multiple comparisons (GraphPad, San Diego, CA, USA). If a significant difference between the treated groups was found, Dunnett's test was used; p-values <0.05 were considered to indicate statistical significance.
Go to :
RESULTS
Effects of murrayafoline A on VSMC proliferation
To investigate whether murrayafoline A could inhibit VSMC proliferation, a non-radioactive colorimetric WST-1 assay and direct cell counting were performed. PDGF-BB (50 ng/ml) increased VSMC proliferation by about two fold compared with unstimulated VSMCs (Fig. 1A). Murrayafoline A decreased PDGF-BB-stimulated VSMC proliferation in a concentration-dependent manner. Fig. 1B shows that the cell number decreased significantly, to 4.5±1.1×104, 2.9±0.8×104, and 2.2±0.1×104 cells/well, as the concentration of murrayafoline A was increased to 1, 3, and 5 µM, respectively. The number of cells increased significantly after 50 ng/ml PDGF-BB treatment (6.5±1.5×104 cells/well) compared with the unstimulated group (2.6±0.8×104 cells/well). Treatment with the highest concentration of murrayafoline A (5 µM) for various incubation times (6-48 h) showed no VSMC cytotoxicity in serum-free medium (Fig. 1C), indicating that the anti-proliferative effect of murrayafoline A on VSMCs was not due simply to a cytotoxic effect.
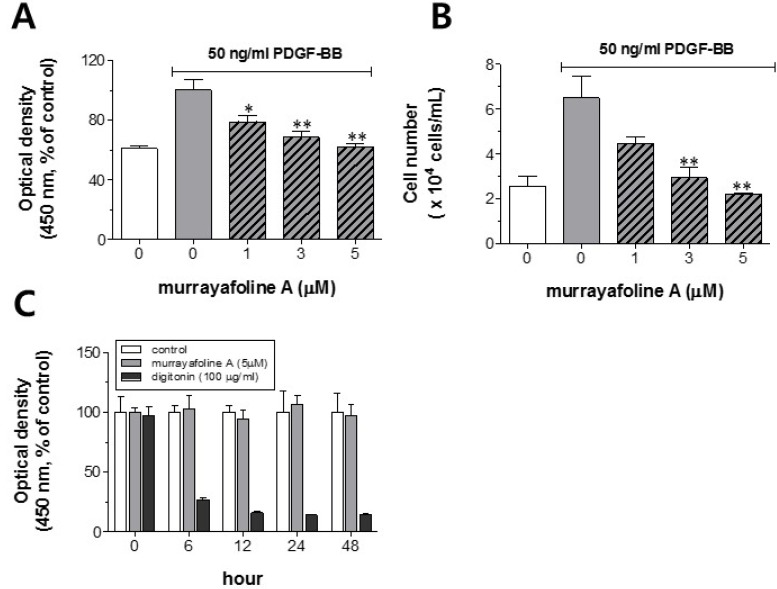 | Fig. 1Effects of murrayafoline A on VSMC proliferation and viability. VSMCs cultured in serum-free medium were stimulated with 50 ng/ml PDGF-BB for 24 h and the effects of various concentrations of murrayafoline A (1-5 µM) on cell proliferation and viability were measured as described in the Experimental Section. (A) Optical densities at 450 nm, as determined in the WST-1 assay (n=3). (B) Cell numbers counted using a hemocytometer (n=4). (C) Viability as determined by the WST-1 assay (n=4) for various incubation times (0-48 h). All values are expressed as means±SEM. Statistical differences from the PDGF-BB control (PDGF-BB-stimulated, but no murrayafoline A) are indicated by *p<0.05 and **p<0.01.
|
Effects of murrayafoline A on the PDGF-BB-induced activation of receptors and downstream molecules in VSMCs
Since PDGF-BB has been known to activate the PLCγ1, protein kinase B (Akt/PKB), ERK1/2, and STAT3 pathways [142021], we examined the inhibitory effects of murrayafoline A on the levels of phosphorylated PDGF receptor β (PDGF-Rβ) and mitogens downstream of PDGF-Rβ signaling pathways. Interestingly, pretreatment of the cells with murrayafoline A did not show any detectable effect on PDGF-Rβ, PLCγ1, Akt, ERK1/2, and STAT3 in PDGF-BB-stimulated VSMCs (Fig. 2). Therefore, these results suggest that the PDGF-Rβ, PLCγ1, Akt, ERK1/2, and STAT3 pathways are not involved in the murrayafoline A-induced inhibition of PDGF-BB-stimulated VSMC proliferation.
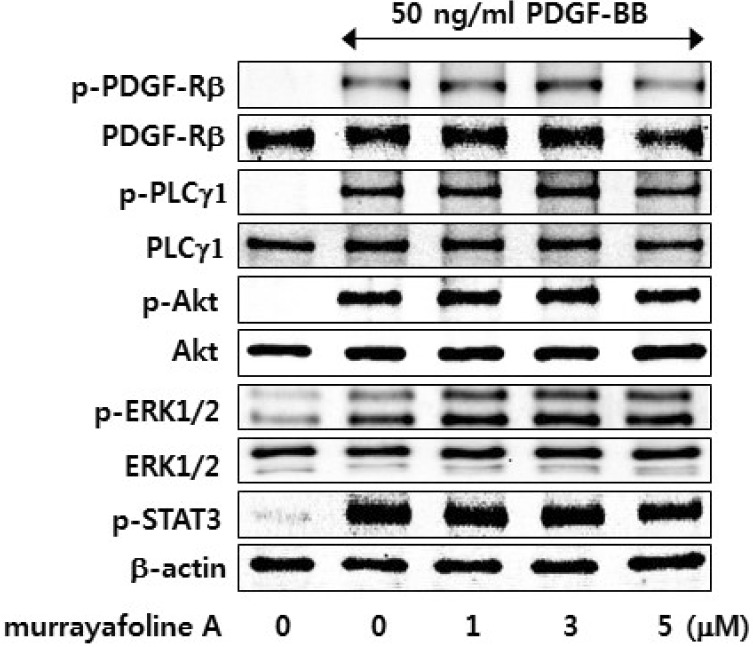 | Fig. 2Effects of murrayafoline A on the PDGF-BB-stimulated activation of PDGF-Rβ, PLCγ1, Akt, ERK1/2, and STAT3. Quiescent VSMCs cultured in serum-free medium were stimulated with 50 ng/ml PDGF-BB, and the effects of various concentrations of murrayafoline A (1-5 µM) in changing the PDGF-BB-induced phosphorylation of PDGF-Rβ, PLCγ1, Akt, ERK1/2, and STAT3 were measured as described in the Experimental Section.
|
Effects of murrayafoline A on DNA synthesis and G0/G1 phase arrest in PDGF-BB-stimulated VSMCs
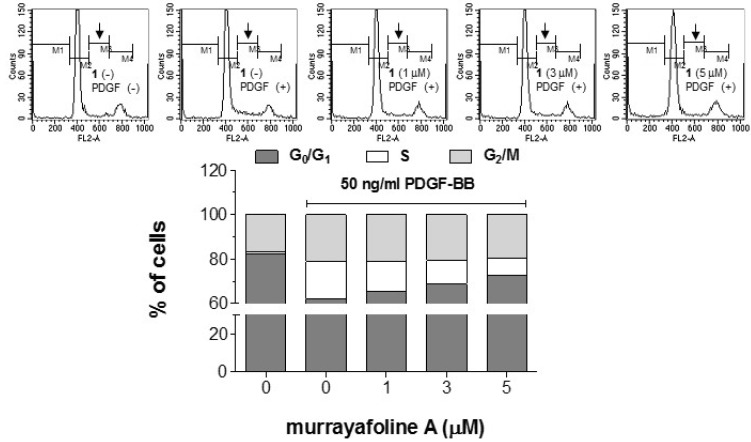
To examine the effects of murrayafoline A on DNA synthesis, a [3H]-thymidine incorporation assay was performed in PDGF-BB-stimulated VSMCs. Fig. 3 indicates that the increased [3H]-thymidine incorporation into DNA caused by PDGF-BB treatment was inhibited significantly by murrayafoline A in a concentration-dependent manner. Representative results of cell cycle analyses showed that the cell population in G0/G1 phase of the cell cycle was increased as the concentration of murrayafoline A was increased in PDGF-BB-stimulated VSMCs (Fig. 4). These results indicate that murrayafoline A induced an arrest at G0/G1 phase of the cell cycle and inhibited progression of the cell cycle to later (S, G2, and M) phases for the proliferation of VSMCs.
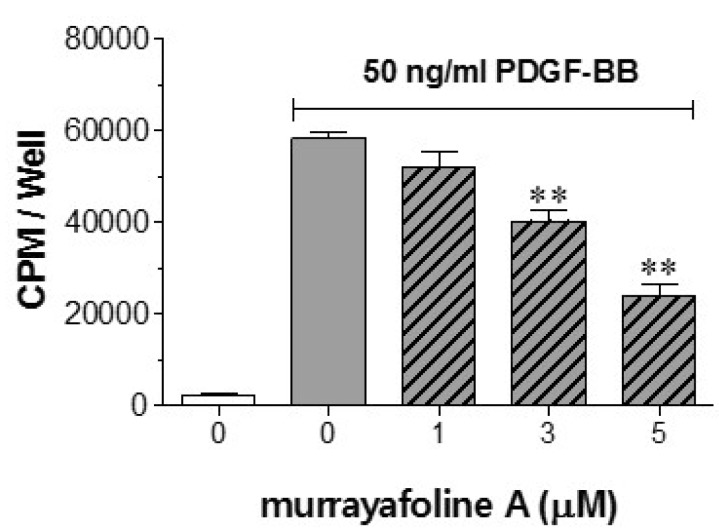 | Fig. 3Effects of murrayafoline A on DNA synthesis. VSMCs cultured in serum-starved medium were stimulated with 50 ng/ml PDGF-BB, and the effects of various concentrations of murrayafoline A (1-5 µM) on [3H]-thymidine incorporation were measured after the addition of 2 µCi/ml [3H]-thymidine, as described in the Experimental Section. The values are expressed as means±SEM (n=4), and statistical differences from the PDGF-BB control (PDGF-BB-stimulated, but no murrayafoline A) are indicated by **p<0.01.
|
Effects of murrayafoline A on the expression and activation of cell cycle-related molecules
It has been known that the cells reach a restriction (check) point in late G1 phase [22]. Beyond this point, the cells are committed to DNA replication, and further cell cycle progression proceeds independently of growth factor stimulation. pRb is a key component of the molecular network controlling this restriction point. Although several CDKs are known to phosphorylate pRb, suppression of CDK2 alone may be sufficient to prevent pRb hyper-phosphorylation [2324]. Hypo-phosphorylated pRb binds to E2F family transcription factors, and thus inhibits the transcription of E2F-responsive genes necessary for cell cycle progression. To examine the underlying mechanism of the murrayafoline A-induced cell cycle arrest, we measured the expression of cyclin D1, cyclin E, CDK2, and CDK4 using immunoblotting. Fig. 5 shows that murrayafoline A significantly inhibited the PDGF-BB-induced expression of cyclin D1/E and CDK2/4 significantly and in a concentration-dependent manner. Moreover, murrayafoline A induced the concentration-dependent inhibition of PDGF-BB-induced pRb hyper-phosphorylation. The expression of proliferating cell nuclear antigen (PCNA), synthesized as a phospho-pRb-mediated gene product in early G0/G1 and S phase of the cell cycle [25], was also inhibited by murrayafoline A in the same pattern as shown for the inhibition of pRb phosphorylation. Taken together, this observation indicates that murrayafoline A inhibits cell cycle progression from G0/G1 to S phase by inhibiting the expression of cyclin D1/E, CDK2/4, and PCNA, and the phosphorylation of pRb in PDGF-BB-stimulated VSMCs.
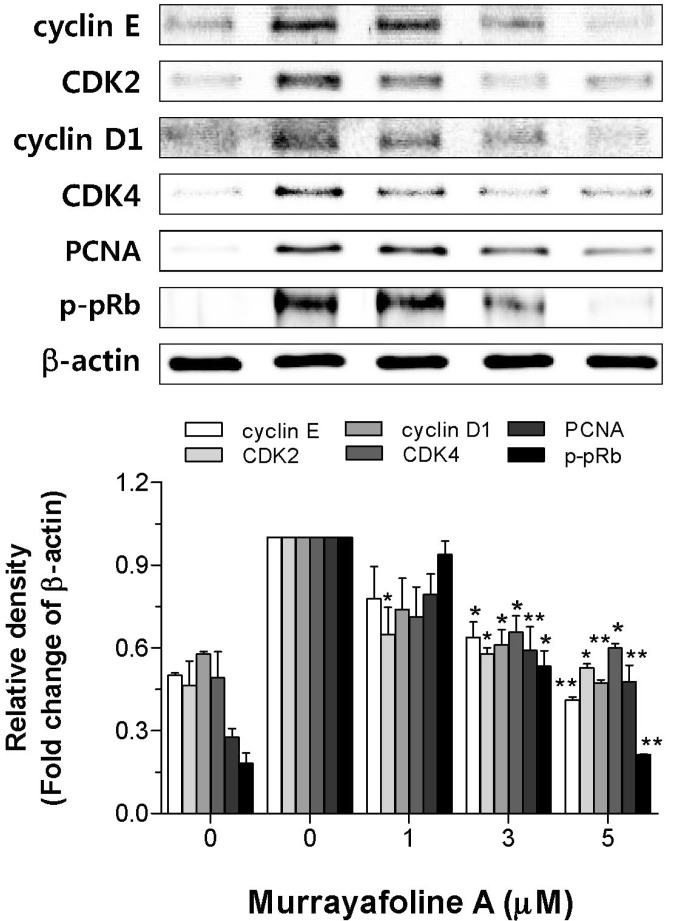 | Fig. 5Effects of murrayafoline A on the inhibition of cell cycle regulatory proteins. Quiescent VSMCs cultured in serum-free medium were stimulated with PDGF-BB to express cell cycle regulatory proteins, and the effects of murrayafoline A on the expression of cyclin E, CDK2, cyclin D1, CDK4, and PCNA, and activation of pRb were assessed as described in the Experimental Section. β-Actin was used for normalization. Immunoblots were analyzed by densitometry and the values are given based on the control of 1.0. The results are an average of four similar experiments, expressed as means±SEM. The insets display representative blots of four similar independent experiments. Statistical differences from the PDGF-BB control (PDGF-BB-stimulated, but no murrayafoline A) are indicated by *p<0.05 or **p<0.01.
|
Go to :
DISCUSSION
This study has two major findings: (1) murrayafoline A has an inhibitory effect on PDGF-BB-induced VSMC proliferation, and (2) this inhibitory effect of murrayafoline A is mediated via an arrest in G0/G1 phase of the cell cycle by suppressing the expression of cyclin-CDK complexes and PCNA, and the pRb phosphorylation in PDGF-BB-stimulated VSMCs. This is the first report of an anti-proliferative action of murrayafoline A in VSMCs. These results indicate that murrayafoline A may be a good candidate for the management of and protection from atherosclerosis and vascular restenosis.
Because the abnormal migration and proliferation of VSMCs in arterial walls are important pathogenic factors in vascular disorders, including atherosclerosis and restenosis after angioplasty [26], the inhibition of VSMC proliferation is a potentially important therapeutic strategy for treating these diseases [27]. In the present study, murrayafoline A showed an inhibitory effect on PDGF-BB-induced VSMC proliferation and this effect was not due to cellular toxicity or apoptosis (Fig. 1). Further investigation indicated that murrayafoline A inhibited DNA synthesis through a G0/G1 cell cycle arrest (Fig. 3, 4). It has been reported that VSMCs are stimulated to divide in response to mitogens after vascular injury, which results in their exit from G1 phase and entry into S phase [28]. Cyclin D1/E and cyclin 2/4 are known to play a positive role in G0/G1 phase and p27 is a negative regulator of CDK/cyclin E complexes, causing a G0/G1 arrest [2930]. Treatment with murrayafoline A resulted in the dose-dependent inhibition of expression of CDK2, CDK4, cyclin D1, and cyclin E (Fig. 5). Although several CDKs are known to phosphorylate pRb, suppression of CDK2 alone may be sufficient to prevent pRb hyper-phosphorylation [2324]. The inhibition of CDK2, CDK4, cyclin D1, and cyclin E expression was sufficient to inhibit phosphorylation of pRb (Fig. 5). Moreover, the expression of PCNA, which is synthesized as a phospho-pRb mediated gene product in early G0/G1 and S phase of the cell cycle [25], was also inhibited (Fig. 5). Thus, our results indicate that the anti-proliferative actions of murrayafoline A in PDGF-BB-stimulated VSMCs are mediated through inhibiting the expression of both cyclin D1/CDK4 and cyclin E/CDK2, leading to a G0/G1 cell cycle arrest.
In conclusion, the present study provides evidence that murrayafoline A inhibits PDGF-BB-induced VSMC proliferation by down-regulating the expression of cyclin D1, cyclin E, CDK2, CDK4, and PCNA, and the phosphorylation of pRb, resulting in arrest at the G0/G1 phase of the cell cycle. This observation indicates that murrayafoline A may be useful for preventing the progression of vascular complications such as restenosis after percutaneous transluminal coronary angioplasty and atherosclerosis.
Go to :
 XML Download
XML Download