

 PDF
PDF ePub
ePub Citation
Citation Print
Print


ABBREVIATIONS
HO-1
heme oxygenase-1
HUVECs
human umbilical vein endothelial cells
SnPP
tin protoporphryin-IX
Nrf2
nuclear factor erythroid-derived 2 related factor 2
ARE
antioxidant response element
OX1R
orexin-1 receptor
OX2R
orexin-2 receptor
VEGF
vascular endothelial growth factor
bFGF
basic fibroblast growth factor
EBM-2
endothelial basal medium
EGM-2
endothelial growth medium
ARE
antioxidant response element
SOD1
superoxide dismutase 1
HIF-1α
hypoxia-inducible factor-1α
mTOR
mammalian target of rapamycin
eIF-4E
eukaryotic translation initiation factor 4E
INTRODUCTION
The orexin peptides, orexin-A and orexin-B, are highly excitatory neuropeptide hormones released by the hypothalamus [1]. Activity of orexins is mediated via two G-protein coupled receptors, orexin-1 receptor (OX1R) and orexin-2 receptor (OX2R) [2]. The orexin system was initially characterized as important in regulation of food intake, sleep/wakefulness, and energy expenditure [3]. More recent studies have reported more diverse physiological actions of orexin-A, such as central and peripheral effects on cardiovascular function [45]. Moreover, we have previously shown that orexin-A induces angiogenesis via the orexin receptors [6], indicating that orexin-A and its receptors play important roles in angiogenesis under pathophysiological conditions.
Angiogenesis is the formation of new capillaries from pre-existing blood vessels, a critical step in physiologic and pathologic events such as embryo development, wound healing, chronic inflammation, and tumor growth [7]. Angiogenesis is regulated by various angiogenic factors, including classic factors such as vascular endothelial growth factor (VEGF) and basic fibroblast growth factor (bFGF), as well as nonclassic endogenous peptides [89].
HO-1 catalyzes the degradation of heme to iron, carbon monoxide, and biliverdin [10]. HO-1 is an anti-inflammatory, antioxidant, and cytoprotective enzyme, which is highly inducible in many different types of cells by a variety of stimuli including heme, heavy metals, hydrogen peroxide, cytokines, and growth factors [11]. Several studies suggest that induction of HO-1 in the vasculature exerts beneficial effects on vascular injury, and mediates protection of endothelial cells, by regulating endothelial cell activation, preventing adhesion molecule expression, inhibiting endothelial cell apoptosis, and promoting formation of new blood vessels [1213141516]. Recent studies have shown a significant role of HO-1 in VEGF- or stromal cell derived factor-1 (SDF-1)-mediated angiogenesis for vascular repair and remodeling [17].
To our knowledge, the present study is the first to show that orexin-A regulates HO-1 expression and enzyme activity in endothelial cells, thereby elucidating the underlying molecular mechanism. We further showed that HO-1 is involved in orexin-A-driven endothelial angiogenesis.
METHODS
Reagents
Orexin-A was purchased from tocris bioscience (Bristol, UK). SnPP was obtained from Enzo Life Sciences (Farmingdale, NY, USA). Human HO-1 antibody and α-tubulin antibody were purchased from Enzo Life Sciences (Farmingdale, NY, USA) and Biogenex (Fremont, CA, USA), respectively. Nrf2 antibody was obtained from Santa Cruz Biotechnology (Santa Cruz, CA, USA). All other chemicals were provided by Sigma-Aldrich (St. Louis, MO, USA), unless otherwise indicated.
Cell culture
HUVECs were purchased from Lonza (Basel, Switzerland) and used at passages 5~8 in all experiments. HUVECs were plated onto 0.2% gelatin-coated dishes and grown in sterile endothelial growth medium (EGM-2 BulletKit, Lonza), composed of endothelial basal medium (EBM-2), trace elements, growth factors, and antibiotics. Human microvascular endothelial cells (HMECs) were obtained from the Center for Disease Control (CDC, USA). HMECs were grown in MCDB 131 medium supplemented with 10% FBS (Gibco BRL, NY, USA), 1% antibiotics, 1 µg/mL hydrocortisone, and 10 ng/mL hEGF (Millipore co., Billerica, MA, USA). Cells were maintained at 37℃ in a humidified mixture of air (95%) and CO2 (5%).
RNA isolation and real-time PCR analysis
Total RNA from HUVECs was isolated using the TRIzol reagent (Invitrogen, Carlsbad, CA, USA) and converted to cDNA using a commercial reverse transcriptase kit (Promega, Madison, WI, USA). Real-time PCR was performed using power SYBR Green (Applied Biosystems, Foster City, CA, USA) with the 7500 Real-Time PCR System (Applied Biosystems, Foster City, CA, USA). HO-1 gene expression was subjected to relative quantification using β-actin for normalization. The following oligonucleotide primers were used for HO-1: 5'-TGAGGAACTTTCAGAAGGGCC-3' and 5'-TGTTGCGCTCAATCTCCTCC-3'; for β-actin the primers used were 5'-ACTCTTCCAGCCTTCCTTCC-3' and 5'-TGTTGGCGTACAGGTCTTTG-3'.
Western blot analysis
HUVECs were lysed with lysis buffer (100 mM Tris-Cl, pH 7.6; 5 mM EDTA; 50 mM NaCl; 50 mM β-glycerophosphate; 50 mM NaF; 0.1 mM Na3VO4; 1 mM PMSF; 0.5% NP-40; 1% Triton X-100; and 0.5% sodium deoxycholate with protease inhibitor cocktail). Equal amounts of protein were separated by SDS-polyacrylamide gel electrophoresis and blotted onto a nitrocellulose membrane (GE Healthcare, Piscataway, NJ, USA). The membranes were probed with appropriate antibodies and the signals were visualized using the enhanced chemiluminescence (ECL) detection system (Thermo Fisher Scientific, Rockford, IL, USA).
HO-1 activity
HO-1 enzyme activity in HUVEC microsomes was measured as rate of bilirubin generation, as described previously [18]. In brief, harvested cells were lysed using MgCl2 (2 mM) phosphate (100 mM) buffer (pH 7.4) with three cycles of freezing and thawing, and then centrifuged at 12,000 g for 15 min at 4℃. The supernatant was incubated with 200 µL reaction buffer (0.8 mM NADPH; 2 mg mouse liver cytosol; 10 µM hemin; 2 mM glucose-6-phosphate and 0.2 U glucose-6-phosphate dehydrogenase) in the dark for 1 hr at 37℃. The formed bilirubin was extracted with 200 µL chloroform, and bilirubin concentration was calculated by the difference in absorbance between 464 and 530 nm (extinction coefficient of 40 mM-1 cm-1 for bilirubin). HO activity was measured as picomole of bilirubin formed/mg of endothelial cell protein/hr.
In vivo Matrigel plug assay
Seven-week-old C57BL/6 mice (Koatech, Pyeongtaek, Korea) were injected subcutaneously with 0.5 mL of Matrigel (BD Biosciences, San Jose, CA, USA) containing orexin-A, SnPP, and heparin (20 units). After 7 days, blood vessel infiltration level in Matrigel plugs was measured by hemoglobin analysis using the Drabkin method. To identify infiltrating endothelial cells, each Matrigel plug was stained with hematoxylin-eosin (H&E) and immunostained with anti-PECAM1 antibody (BD Biosciences, San Jose, CA, USA), a marker for endothelial cells.
Aortic ring assay
Aorta were harvested from 6-week-old Sprague-Dawley rats (Koatech, Pyeongtaek, Korea) and sectioned into 1-mm-thick slices (aortic rings). The rings were placed on wells coated with Matrigel (50 µL). The rings were incubated at 37℃ in 5% CO2 for 72 hr, with or without orexin-A and SnPP. Each aortic ring was photographed under a phase contrast microscope and microvessel outgrowth was quantified by counting the number of sprouts.
Tube formation assay
HUVECs were plated onto a 24-well plate coated with 300 µL growth factor-reduced Matrigel (BD Biosciences, San Jose, CA, USA). Cells were incubated in M199 culture medium (Gibco BRL, NY, USA) containing 1% FBS, with or without orexin-A and SnPP, for 4 hr at 37℃. Morphological changes of the cells were observed under an inverted microscope and photographed. Tube formation was quantified by measuring the tube areas.
Chemotactic migration assay
Transwell membranes (8 µm pore size; Corning Costar, Cambridge, MA, USA) were coated on the lower surface of the filter with 10 µg of gelatin. Six hundred µL of M199 medium (1% FBS), with or without orexin-A and SnPP, was placed in the lower chamber. HUVECs were suspended at a final concentration of 105 cells per 100 µL in M199 medium containing 1% FBS. The cell suspension was loaded into each of the upper wells and incubated at 37℃ in 5% CO2 for 4 hr. HUVECs that migrated to the lower surface were fixed and stained with H&E. The number of migrated cells was counted under an optical microscope.
Transient transfection and reporter gene analysis
HMECs in 24-well plates were transiently transfected with the pGL3-ARE-Luc promoter luciferase construct and pCMV-β-gal using X-tremeGENE (Roche, Madison, WI, USA). ARE-Luc reporter plasmid was a generous gift from Dr. Park, R. K. (Wonkwang University, Korea). After 21 hr, cells were incubated with orexin-A for a further 3 hr. Cell extracts were prepared, and analyzed for luciferase activity using a luciferase assay system (Promega, Madison, WI, USA) and a luminometer (Turner Biosystems, Sunnyvale, CA, USA). Luciferase activity was normalized against the co-transfected β-galactosidase enzyme activity. Assays were repeated at least three times.
RNA interference
Nrf2 siRNA and control siRNA were purchased from Santa Cruz Biotechnology (#sc-37030, Santa Cruz, CA, USA) and Bioneer (#SP-1013, Daejeon, Korea), respectively. Nrf2 knockdown of HUVECs was performed using the HUVEC kit and Amaxa nucleofector (Lonza, Basel, Switzerland). Transfected cells were seeded in a 35 mm dish. After incubation, cells were treated with orexin-A, and a Western blot analysis and tube formation assay was performed.
RESULTS
Orexin-A induces HO-1 expression and activity in human vascular endothelial cells
To examine whether orexin-A affects HO-1 expression in vascular endothelial cells, HUVECs were treated with orexin-A and whole cell lysates were analyzed by Western blot. As shown in Fig. 1A, a time-dependent increase in HO-1 protein expression was observed in endothelial cells after orexin-A treatment. Furthermore, orexin-A induced HO-1 expression in a dose-dependent manner (Fig. 1B). Real-time PCR demonstrated that expression of HO-1 mRNA was also induced by orexin-A (Fig. 1C). By contrast, there was no effect on HO-2 mRNA levels in HUVECs incubated with orexin-A (Fig. 1D). An increase in HO-1 enzyme activity was also observed after orexin-A treatment, as determined by in vitro bilirubin production, and this increase was effectively attenuated by SnPP, a HO-1 inhibitor (Fig. 1E). These results indicate that orexin-A induces HO-1 mRNA, protein, and enzyme activity in vascular endothelial cells.
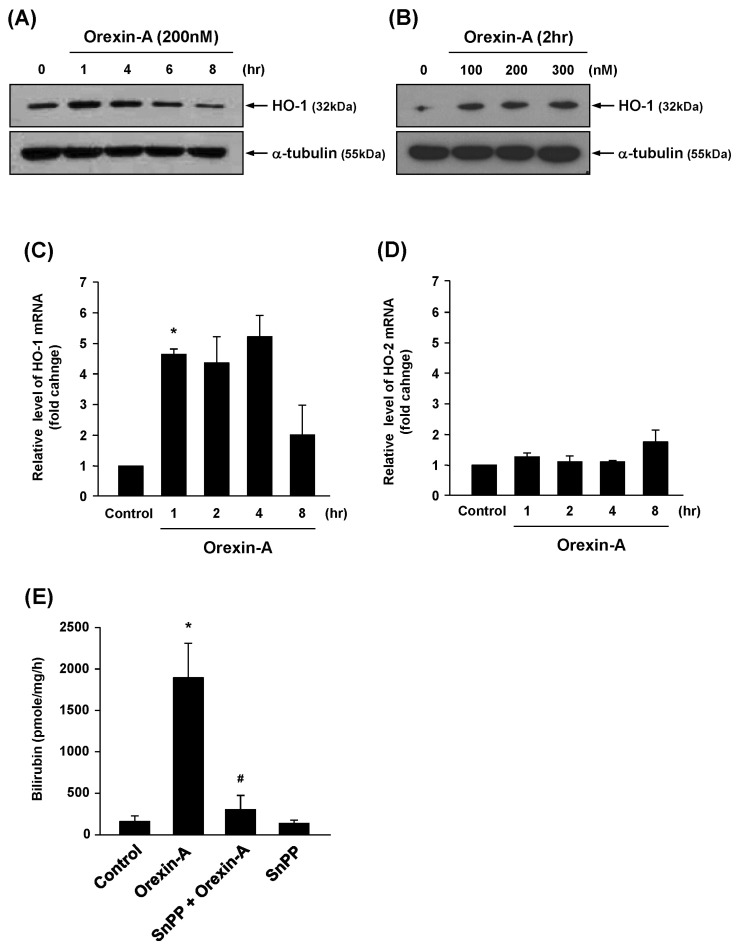
Fig. 1
Upregulation of HO-1 expression and activity by orexin-A in vascular endothelial cells. (A and B) HUVECs were treated with orexin-A (200 nM) or phosphate buffered saline (PBS) for the various times and concentrations shown. Expression of HO-1 was determined by western blot using HO-1 and α-tubulin antibodies (loading control). (C and D) Cells were incubated with orexin-A (200 nM) or PBS for the indicated times. Expression of HO-1 and HO-2 mRNA was analyzed by real-time PCR. These data represent the mean±SE of three experiments. *p<0.05 vs. control. (E) HUVECs were pretreated with SnPP (50 µM) for 15 min, followed by treatment with orexin-A for 4 hr. HO-1 enzyme activity was measured as described in the Materials and Methods. Three independent experiments were performed. *p<0.05 vs. control; #p<0.05 vs. orexin-A alone.
![]()
HO-1 plays a role in orexin-A-induced angiogenesis in vivo
Previously, we have suggested that orexin-A exerts angiogenic properties both in vivo and ex vivo [6]. To further assess whether increased HO-1 expression and activity is required for orexin-A-induced angiogenesis, Matrigel containing orexin-A, with or without SnPP, was subcutaneously injected into male C57BL/6 mice. After 7 days, the Matrigel plugs from each mouse were collected and analyzed. As shown in Fig. 2A, Matrigel containing orexin-A showed a remarkably reduced angiogenic effect in the presence of SnPP when compared with Matrigel containing orexin-A alone. The degree of blood vessel formation in the Matrigel plug was confirmed by measuring hemoglobin content (Fig. 2B). In addition, histological analysis indicated that inhibition of HO-1 by SnPP resulted in decreased levels of orexin-A-induced infiltration of endothelial cells (Fig. 2C). Infiltrated cells were positive for the endothelial marker PECAM-1 (Fig. 2D). These data demonstrate that HO-1 activity is required for orexin-A-mediated angiogenesis in vivo.

Fig. 2
Involvement of HO-1 in orexin-A-induced angiogenesis in vivo. (A) Matrigel was treated with orexin-A (200 nM) or PBS in the presence or absence of SnPP (20 µM), and then subcutaneously injected into mice. After 7 days, the plugs were obtained from each mouse and photographed. (B) The level of vessel formation was quantified by measuring hemoglobin content. Each value represents the mean of at least three animals. *p<0.05 vs. control; #p<0.05 vs. orexin-A alone. (C) Matrigel plugs were stained using H&E and photographed. (D) Endothelial cells recruited into Matrigel plugs were immunostained with PECAM-1 antibody and observed using fluorescence microscopy.
![]()
HO-1 activation regulates the angiogenic activity of orexin-A in vascular endothelial cells
To assess the involvement of HO-1 in orexin-A-induced endothelial angiogenesis ex vivo, the aortic ring angiogenesis assay was used. As shown in Fig. 3A, orexin-A promoted endothelial cell sprouting from aortic segments, whereas the presence of SnPP resulted in a significant decrease in orexin-A-stimulated microvessel sprouting. The role of HO-1 in orexin-A-driven angiogenesis was further assessed by measuring the ability of HUVECs to form capillary-like networks. Orexin-A stimulated tubular formation by vascular endothelial cells on Matrigel. Co-treatment of HUVECs with orexin-A and SnPP resulted in a reduction in orexin-A-induced endothelial tube formation (Fig. 3B, left). Tube-like structures formed by HUVECs were quantified by measuring the number of tube areas (Fig. 3B, right). Next, we investigated the influence of HO-1 on the chemotactic motility of HUVECs using a modified Boyden chamber assay. As shown in Fig. 3C, orexin-A stimulated the migration of HUVECs, and substantial inhibition of orexin-A-mediated migration of HUVECs was observed in the presence of SnPP. Collectively, these results show that HO-1 mediates orexin-A-induced endothelial angiogenesis ex vivo and in vitro.

Fig. 3
Effect of HO-1 inhibition on orexin-A-stimulated angiogenesis ex vivo and in vitro. (A) Rat aortic rings were embedded in Matrigel and cultured with orexin-A (200 nM) for 4 days in the presence or absence of SnPP (20 µM). The sprouted microvessels from aortic rings were photographed under a microscope (left). The newly formed sprouts of endothelial cells were counted (right). (B) HUVECs were seeded on growth factor-reduced Matrigel in the presence or absence of SnPP (20 µM) and treated with orexin-A (200 nM) for 4 hr. Cells were observed under a phase contrast microscope and the number of tube areas was counted. (C) HUVECs were seeded on gelatin-coated filters of transwell chambers. Cells were incubated with orexin-A (200 nM) and SnPP (20 µM) for 4 hr. The filter containing migrated cells was stained with H&E and photographed. The number of migrated cells was counted. All results shown are representative of at least three independent experiments. *p<0.05 vs. control; #p<0.05 vs. orexin-A alone.
![]()
Orexin-A-induced expression of HO-1 and angiogenic activity is associated with activation of Nrf2
Nrf2 is a transcription factor for antioxidative genes and their proteins, including HO-1. Nrf2 induces HO-1 by binding to a cis-acting enhancer element known as the ARE. In order to test the influence of orexin-A on ARE promoter activity, we performed a luciferase reporter gene assay. Vascular endothelial cells were transiently transfected with the ARE-luciferase reporter gene, and luciferase activity was determined after orexin-A treatment. As shown in Fig. 4A, ARE-luciferase activity was increased in orexin-A treated cells. To further determine whether Nrf2 is involved in orexin-A-induced HO-1 expression, we transiently transfected endothelial cells with Nrf2 siRNA or control siRNA. Silencing with Nrf2 siRNA decreased basal levels of Nrf2 and HO-1 protein expression compared with those in control siRNA-transfected cells (Supplemental Fig. 1). The induction of Nrf2 and HO-1 protein by orexin-A was diminished in Nrf2 siRNA-transfected cells when compared with control siRNA-transfected cells (Fig. 4B). Furthermore, as shown in Fig. 4C, knockdown of Nrf2 also decreased angiogenic activity of orexin-A in a tube formation assay. These data suggest that Nrf2-ARE is closely correlated with orexin-A-mediated angiogenesis through HO-1.
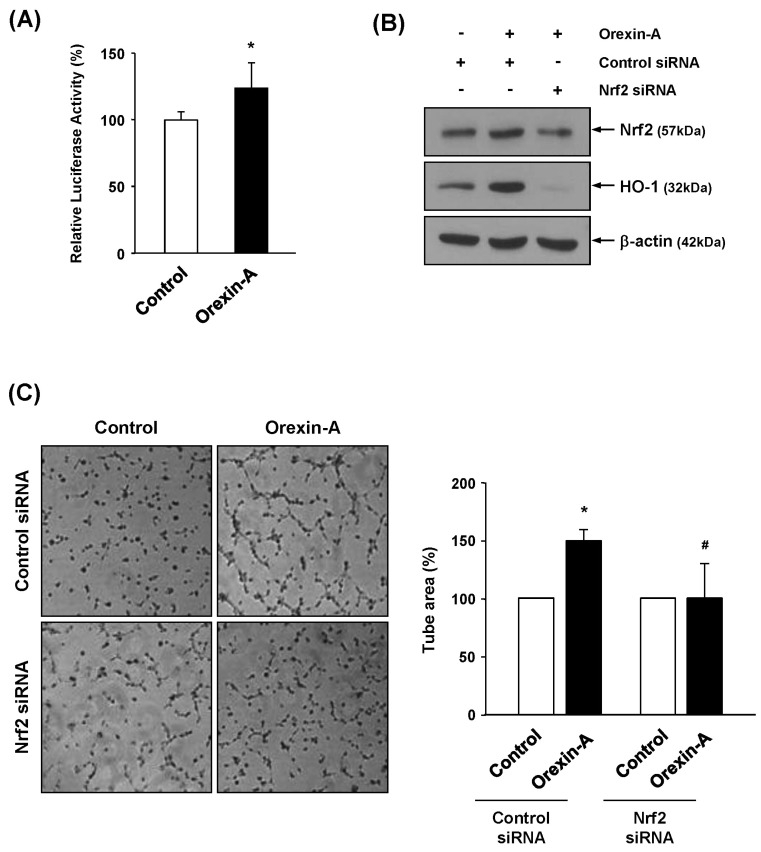
Fig. 4
Influence of the ARE-Nrf2 pathway in orexin-A-mediated HO-1 induction and angiogenesis. (A) HMECs transfected with an ARE-luciferase construct were treated with orexin-A (200 nM) for 3 hr. The cell extracts were prepared and analyzed using a luminometer. Three independent experiments were performed. *p<0.05 vs. control. (B) HUVECs were transiently transfected with control or Nrf2 siRNA (1 µM) by Amaxa nucleofector, followed by treatment with orexin-A for 2 hr. The expression of Nrf2 and HO-1 was determined by Western blot analysis. (C) For tube formation assays, Nrf2 siRNA-transfected cells were seeded on growth factor-reduced Matrigel. Cells were incubated with orexin-A (200 nM) for 4 hr. Newly formed tubes were photographed, and tube areas were quantified from at least three individual experiments. *p<0.05 vs. control; #p<0.05 vs. control siRNA with orexin-A.
![]()
DISCUSSION
Orexin-A and B were first identified as neuropeptides secreted from neurons within the lateral hypothalamus, and were characterized as central regulators of sleep-wake cycles, energy homeostasis, and feeding behavior [319]. Recently, an increasing body of evidence suggests that orexins and orexin receptors, OX1R and OX2R, are present in a variety of peripheral tissues [20]. Expression of prepro-orexin, the precursor peptide of orexin-A and orexin-B, has been reported in peripheral tissues. Interestingly, there is evidence that orexin-A, and not orexin-B, can cross the blood-brain barrier [21]. Plasma orexin-A is present in human and rat peripheral circulation, although the major source of plasma orexin-A remains to be clarified [2223]. Orexin receptors show abundant expression in many peripheral tissues, and mediate functional responses when exposed to exogenous orexins in a wide range of organs outside the central nervous system, such as the gastrointestinal tract, adrenal gland, pancreas, and adipose tissue [20]. We previously found the expression of orexin receptors, OX1R and OX2R, in vascular endothelial cells [6], and it has also been shown that exogenous orexin-A induces orexin receptor-mediated endothelial angiogenesis [6]. However, for other peripheral tissues, the function of orexin and its mechanisms of action are currently not known.
Recent studies have reported that orexin-A exerts a neuroprotective effect in cerebral ischemia and in a cellular model of Parkinson's disease [2425]. Orexin-A was also shown to contribute to the protection of gastric mucosa against acute gastric damage [26]. Cytoprotective mechanisms against noxious stimuli and oxidative stress are mainly mediated by induction of phase 2 enzymes, including HO-1 and superoxide dismutase 1 (SOD1), expression of which is in turn regulated by transcription factor Nrf2 [27]. In this study, we confirmed that orexin-A induces the activation of Nrf2, with consequent expression of HO-1. Further investigations are needed to clarify the role of the Nrf2-HO-1 axis and its mechanism of action in orexin-A-induced neuroprotection and gastroprotection.
A growing body of evidence suggests that HO-1 induction could be a significant event in vasculogenesis and angiogenesis [28]. Among the different end-products generated by HO-1 induction, CO is proposed to be involved in regulating vessel formation, by inducing the production of angiogenic growth factors, and by stimulating endothelial cell/endothelial progenitor cell proliferation and migration by release of VEGF and SDF-1 [29]. CO increases the production of VEGF by stabilization of hypoxia-inducible factor-1α (HIF-1α) protein levels [30]; HIF-1α is active in orexin-A-stimulated cells even under normoxic conditions [31]. Additional studies are needed to investigate whether the HO-1/CO pathway mediates orexin-A-induced HIF-1α activity in endothelial cells.
In addition, we observed that treatment with actinomycin D, a transcriptional inhibitor, did not significantly regulate orexin-A-induced HO-1 protein expression. However, cycloheximide, an inhibitor of protein translation, blocked HO-1 protein expression induced by orexin-A (Supplemental Fig. 2). These results suggest that orexin-A-mediated HO-1 induction occurs predominantly at the translational level. Moreover, orexin-A has been recently reported to stimulate the activation of mammalian target of rapamycin (mTOR) [32]. mTOR regulates mRNA translation by phosphorylation of the eukaryotic translation initiation factor 4E (eIF-4E) binding proteins and the S6 kinases [3334]. Thus, the possibility is raised that orexin-A may increase HO-1 expression via activation of the mTOR downstream target molecules including translational regulatory proteins. This possibility is under investigation.
In this report, we showed that HO-1 mediates orexin-A-induced chemotactic migration and tube formation of endothelial cells. Previous studies have shown that proangiogenic factors such as VEGF and SDF-1 upregulate HO-1 expression in endothelial cells [3517]. Therefore, we cannot exclude the possibility that VEGF or SDF-1 contribute to the proangiogenic effects of orexin-A and HO-1 gene expression induced by orexin-A.
In conclusion, the present study demonstrates orexin-A-stimulated neovascularization to be closely linked to Nrf2-mediated HO-1 expression in endothelial cells. Additional in vivo studies are needed to assess whether the orexin-A-HO-1 axis represents a possible therapeutic target in the management of angiogenesis-related diseases.
 XML Download
XML Download