

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Bone is a very complicated tissue in which formation and resorption occur by a bone remodeling process. The loss of bone mass occurs in osteoporosis. Due to low bone mass, there is increased bone fracture of hip, shoulder, wrist and spine. Estrogen is the key hormone for maintaining bone mass. It has been thought that estrogen deficiency is the major cause of age-related bone loss in humans [1]. Osteoporosis is initiated by the failure to acquire the optimal peak of bone mass during growth and/or to maintain bone mass in later years [2]. The rapid loss of bone mass occurs in post-menopausal women, mainly due to the net decrease of estrogen [2]. The estrogen deficiency also induces an imbalance in bone remodeling, accelerates the loss of bone mass increasing fracture risk [34]. Therefore, estrogen deficiency can be characterized by high bone turnover with enhanced osteoclastic bone resorption. The important factor in determining bone density and bone turnover in humans is circulating estradiol [56789]. The most abundant estrogen precursor in circulation is the sulfate conjugate form of estrogen such as estrone sulfate. The amount of this conjugated form of estrogen is several fold higher than that of unconjugated estrogens [101112]. In addition, the estrone sulfate is well known as the substrate of organic solute carriers such as SLC22A8 (hOAT3), SLC22A9 (hOAT7) and SLC22A11 (hOAT4) [131415].
The single non-synonymous SNPs (nsSNPs) contribute commonly to the functional diversity on encoded proteins in humans. It appears that nsSNP often lead to unexpected responses to drugs and changes in the susceptibility to disease [16]. A number of reports indicated that nsSNPs affect the functional roles of certain proteins in gene regulation by altering DNA and transcription factor binding and/or by affecting signal transduction [171819].
In this report we screened SNPs of SLC22A9 in two groups of Korean women: one group had osteoporosis and the second was a normal control group. We then performed a functional analysis of the nsSNP from the osteoporosis group by expression in Xenopus oocytes.
METHODS
Materials
Taq DNA polymerase and polymerase chain reaction (PCR) primers were purchased from ELPIS BIO (Daejeon, Korea) and Bioneer (Daejeon, Korea), respectively. Restriction enzymes, oligo (dT), LA Taq DNA polymerase and dNTPs were bought from TaKaRa Korea Biomedical Co. (Seoul, Korea), and rNTP and m7G(5')ppp(5')G Cap analogue for synthesis of cRNA were purchased from Amersham Pharmarcia Biotech (Sunnyvale, CA, USA). T7 RNA polymerase was bought from Stratagene Cloning Systems Inc. (La Jolla, CA, USA) and RNasin (RNA inhibitor) was purchased from Promega (Madison, WI, USA). The DNA blood mini kit was purchased from Qiagen (Hilden, Germany). [3H]estrone sulfate (53 Ci/mmol) was purchased from NEN Life Science Products, Inc. All other chemicals utilized in this study were insured for the highest purity available from commercial sources.
Patients
Blood samples were taken from 50 osteoporosis patients and 50 normal control subjects, which were approved by the Samsung Hospital Institutional Review Board (Cheil General Hospital & Women's Healthcare Center IRB). All subjects involved in the present study were age matched (40 to 65 years of age). The blood samples were kindly supplied from Dr. Ki Ok Han and also employed previous research [20].
PCR reaction
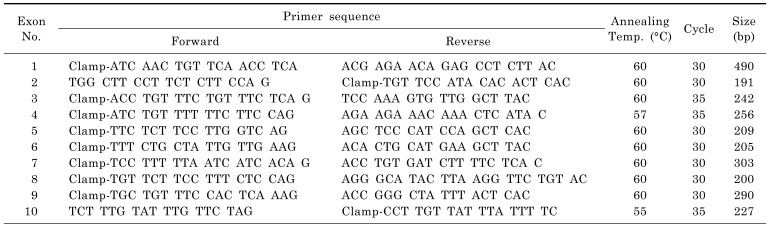
Genomic DNAs were extracted and used as template for the PCR reaction. The PCR primers were approximately 18 mers and were before/after each exon with the GC clamp on the forward or reverse primer [21]. Each PCR reaction mixture (final volume of 12.5 µl) contained 50 ng of isolated genomic DNA, 0.5 mM of dNTP, 10 pmol of each primer and 1 unit of DNA taq polymerase. Amplification was performed using a Takara thermocycler (TP-3000) and the PCR cycling conditions were as follows: an initial denaturation at 94℃ for 2 min, followed by 30 or 35 cycles of denaturation at 94℃ for 1 min, annealing for 1 min at 55℃ to 60℃, and elongation at 72℃ for 1 min. The last cycle was followed by an additional elongation step of 72℃ for 10 min. For optimal DGGE analysis, amplification was followed by a heteroduplexing step, which includes denaturation at 98℃ for 10 min, renaturation for 30 min at the annealing temperature of amplification, and extension at 37℃ for 10 min. The amplified products were checked by electrophoresis using 1.5 µl (about 1% of total volume) of each product in a 1% agarose gel stained with 1 µg/ml of ethidium bromide solution. The primer sequences and annealing temperature are shown in Table 1.
Denaturing gradient gel electrophoresis (DGGE)
DGGE was performed using a V20-HCDC unit (www.scie-plas.com). PCR products were loaded onto a 230×250 mm, 0.5 mm-thick polyacrylamide gel (acrylamide: bisacrylamide, 37.5:1) containing linear denaturing gradients (100% UF=7 M urea/40% deionized formamide) [2223]. The percentage of polyacrylamide was 10% and UF solution gradient was from 10% to 70%. Electrophoresis was performed in TAE buffer (40 mM Tris, 20 mM Acetic acid and 1 mM EDTA) at 250 V for 3.5 h at 60℃. The gels were stained with ethidium bromide solution (1 µg/ml) for 10 min and photographed under an UV transilluminator.
DNA sequencing and variant confirmation
Samples showing aberrant DGGE bending patterns were used in PCR with non-GC clamp primers prior or after 80 base pairs of the designated site. Samples showing additional heteroduplex bands were subjected to sequencing analysis for the exact determination of the sequence variant.
Mutant construction
The wild type SLC22A9 cDNA was obtained by screening a human kidney cDNA library and subcloned to pcDNA 3.1(+). In order to prepare mutants SLC22A9, a modified PCR was performed using primer sets containing both mutation and restriction enzyme sites (Table 2). PCR was sequentially performed for 12 cycles with the first primer (0.5 µM, 1 µl) and 12 cycles with the second primer (0.5 µM, 1 µl) and finally 35 cycles with the third primer (10 µM, 1 µl). The PCR product was sub-cloned using the TA cloning vector (Invitrogen) and confirmed its nucleotide sequences. The mutant constructs were prepared by substituting the mutated product into the wild type cDNA of SLC22A9 using the appropriate restriction enzyme digested cDNA.
cRNA synthesis and uptake experiments using Xenopus laevis oocytes
cRNA synthesis and uptake experiments were performed as described previously [24]. The capped cRNAs were synthesized in vitro with T7 polymerase from plasmid cDNAs linearized with Xho I. Defolliculated oocytes were injected with 25 ng of the capped cRNAs and incubated in Barth's solution (88 mM NaCl, 1 mM KCl, 0.33 mM Ca(NO3)2, 0.4 mM CaCl2, 0.8 mM MgSO4, 2.4 mM NaHCO3, and 10 mM Hepes) containing 50 µg/ml gentamycin and 2.5 mM pyruvate, pH 7.4, at 18℃. After incubation for 2~3 days, uptake experiments were performed at room temperature in ND96 solution (96 mM NaCl, 2 mM KCl, 1.8 mM CaCl2, 1 mM MgCl2, and 5 mM Hepes, pH7.4). The uptake reaction was initiated by replacing ND96 solution containing various concentrations of radio-labeled estrone sulfate, which was terminated by the addition of sufficient ice-cold ND96 solution followed by five times washing after 1 h incubation. Oocytes were solubilized with 10% SDS and the radioactivity was determined by scintillation counting.
RESULTS
Genomic organization of human SLC22A9
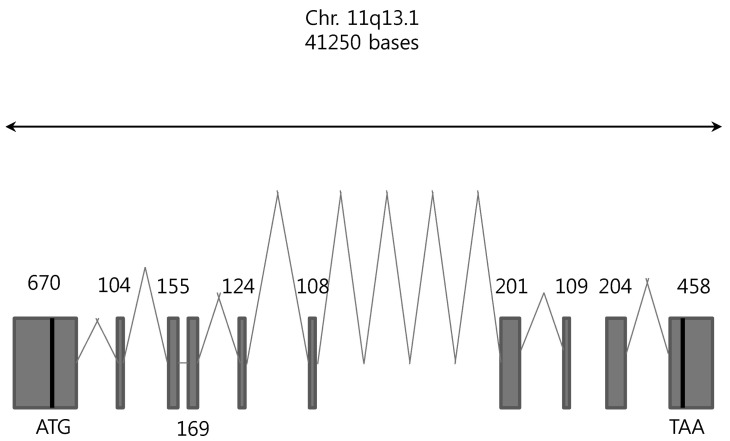
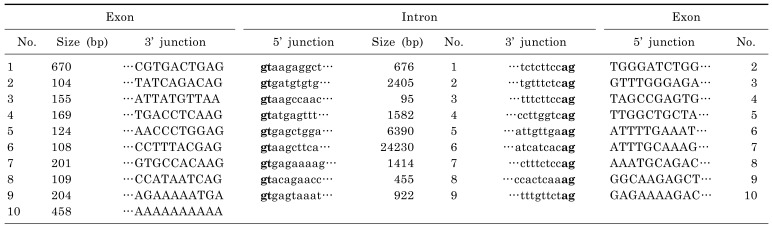
Using the SLC22A9 cDNA nucleotide sequence, we could find the SLC22A9 genomic DNA was from the NCBI database (gi|568815587:63369670-63410919 Homo sapiens chromosome 11, GRCh38 Primary Assembly). The gene, ~41.3 kb long, is located on chromosome 11q13.1. Using the cloned SLC22A9 with the reported genomic sequence, an alignment of the nucleotide sequence was performed to determine the exon-intron gene organization. The SLC22A9 gene consists of 9 introns and 10 exons (Fig. 1). The size of each exon, intron, and nucleotide sequence of the splice junction is shown in Table 3. The consensus sequences for RNA splicing (gt/ag) are found in the 5' and 3' termini for each intron. The translation start codon (ATG) is present in exon 1, and the translation termination codon (TAG) is present in exon 10.
GC-clamp PCR, DGGE assay and screening for sequence variation
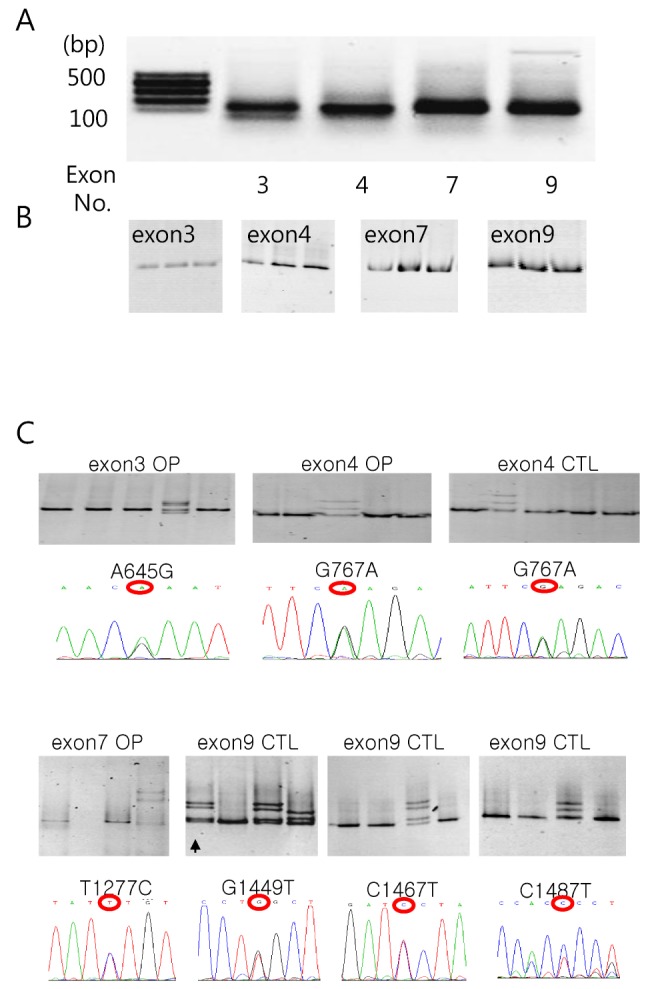
The DGGE method was used in order to determine the SNP. The PCR conditions were tested over several annealing temperatures and cycle numbers using normal human chromosomal DNA. The typical patterns of each PCR products with synonymous and non-synonymous mutations found using GC-clamping primer sets separated on 1% agarose gel are presented in Fig. 2A. The representative results of DGGE separated on acrylamide gel using several control samples are shown in Fig. 2B. The results of DGGE showed a single band in the SNP found in exons.
To determine the SNP in Korean osteoporosis patients, 50 blood samples were obtained from osteoporosis and normal subjects and screened. When the osteoporosis and control subjects were analyzed, six subjects showed different electrophoretic patterns (Fig. 2C). In general, PCR products were produced in accordance with the different number of bands indicating that a mutation may have occurred. Among these six SNPs, two SNPs were found in osteoporosis patients (one in exon 3 and the other in exon 7), three SNPs were found in exon 9 of normal subject and one SNP was found in both osteoporosis patients and normal subjects (exon 4). In order to further clarify this, nucleotide sequencing was performed. As shown in Fig. 2C, the SNPs found from exon 5, 7 and 9 showed that G in the 767th nucleotide of the coding region was changed to A (arginine to glutameine), T in the 1277th nucleotide of the coding region was changed to C (phenylalanine to serine) and C in the 1487th nucleotide of the coding region was changed to T (proline to leucine), respectively. However, other SNPs did not change the amino acid sequences.
Functional analysis of human SLC22A9 variant in Xenopus oocytes
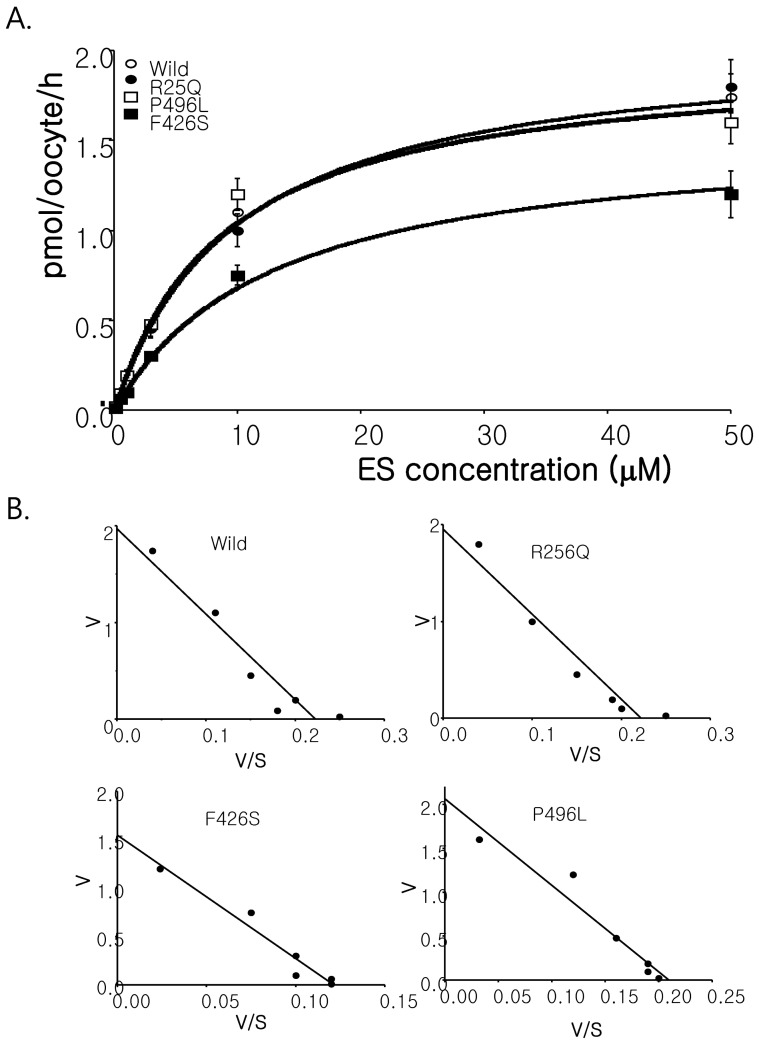
In order to examine whether the SLC22A9 polymorphism found in Korean osteoporosis patients affects the functional activity on substrate transport, we constructed mutant cDNAs and expressed them into Xenopus oocytes. As shown in Fig. 3, F426S exhibited reduced uptake for [3H] estrone sulfate compared with that of the wild-type SLC22A9. R256Q and P496L did not show a statistically significant change in [3H] estrone sulfate compared with those of the wild-type SLC22A9. According to the changes of concentration-dependent [3H] estrone sulfate in wild type and F426S, the Km values for wild-type and F426S were 0.7 and 1.2 µM, respectively, and the Vmax values for wildtype and F426S were 1.8 and 0.47 pmol/oocyte/h, respectively (Fig. 3).
DISCUSSION
In the present study, the genetic feature of the SLC22A9 was investigated in the Korean women osteoporosis patients and normal subjects. We identified six SNPs (A645G, G767A, T1277C, G1449T, C1467T and C1487T) of SLC22A9 coding region from the osteoporosis and control subjects. Totally 3082 SNPs are reported in the National Center for Biotechnology Information (NCBI) data base. Among them, 1,335 synonymous and non-synonymous SNPs are observed in coding regions. They consist of 68 missense mutations and three stop codon gained mutation within the coding region. Two nsSNP, T1277C (F426S) and C1487T (P496L) of SLC22A9, ares not yet reported in the NCBI data base. We also verify the existence of a functional alteration in F426S.
Human SLC22A9, a sulfate conjugate transporter, transports estrone- and dehydroepiandrosterone sulfate as predominant substrates. In two separate studies, Grumbach and Bilezikian showed estrogen plays a major role in the control of bone growth as well as in the accumulation and maintenance of bone mass. This was shown in using a knockout mouse model that contained mutations impairing estrogen action or formation [2526]. Bone mass reaches a peak in adults in their twenties and thirties. It begins to decline from age 35~40. The levels of dehyroepiandrosterone and its sulfate conjugate, precursors of androgens and estrogens, decline 2% per year, leaving a residual of 10~20% of the peak production by the eighth decade of life. The age-associated decrease may lead to osteoporosis. Therefore, it is important to understand steroid or steroid conjugate transporter in HRT.
Numerous large-scale screening were carried out to identify genetic variations that affect disease susceptibility and drug response as well as to determine the variants contributing to alter the phenotype. We found three nsSNPs from 50 Korean women osteoporosis patients and normal controls. Unfortunately, there is no information of any functional analysis with discovered nsSNP. Our functional analysis showed the increased value in Km (wild type, 8.84 µM vs. F426S, 12.74 µM) and decreased value in Vmax (wild type, 1.97 pmol/oocyte/h vs. F426S, 1.55 pmol/oocyte/h) in estrone sulfate transport. The transporting activities of the other two nsSNPs (R256Q and P496L) are similar to those of wild type SLC22A9.
In human, the functional properties of transport have been reported. With respect to substrate specificity, hOAT3 (SLC22A8) and hOAT4 (SLC22A11) also transport estrone and dehyroepiandrosterone sulfate. The differences between hOAT3, hOAT4 and SLC22A9 are the expression pattern in organ. The expression of OAT3 was observed in brain, skeletal muscle and kidney [13]. The OAT4 message was observed in kidney and placenta [15]. In contrast, SLC22A9 was expressed exclusively in liver [14]. The result of our ongoing study using Saos-2 and MG-63 osteosarcoma cell line showed the expression of SLC22A11 (hOAT4) and SLC22A9 (hOAT7) by polymerase chain reaction (data not shown). From these results, it might be thought of as indirect evidence of steroid sulfate conjugate transport in bone. When we screened the SNPs of SLC22A8 using the same patient blood samples, no nsSNP was found. In the case of SLC22A11 (hOAT4), we have found one nsSNP and have reported the functional change of transporting activity of the mutant (E278K) in independent osteoporosis patients [20]. In order to understand the characteristics of SLC22A8, 9 and 11, the study using cells derived from primary cultured or immortalized cells was necessary.
Several transporter proteins such as the multidrug resistance associated protein [2728], organic anion transporting polypeptide C [2930], Na+-taurocholate cotransporting polypeptide [3132] and the breast cancer resistance protein (BCRP/ABCG2) [3334] also transport steroids or their sulfate conjugates and their nsSNPs also showed a decrease of their substrate transporting abilities [35363738]. It has also reported that nsSNPs showing altered activity in some transporters is co-related with certain disease [3940].
In addition, several reports revealed that some SNPs of SLC22A8 and SLC22A11 are located in the promoter site [41]. NCBI data base also shows some upstream SNP. The study on the regulatory effect of upstream SNP will be also necessary.
In conclusion, we reported for the first time the nsSNPs of SLC22A9 in Korean women with osteoporosis and verified altered activity that may contribute to understanding certain diseases including osteoporosis.

 XML Download
XML Download