

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Finding effective anti-aging strategies has recently become one of the most active researches in the life sciences. Calorie restriction (CR) has been well established as the most effective intervention to extend the lifespan and alleviate age-related disorders in mammals, including mice and monkeys. Although CR has not been verified to extend the lifespan in humans [1,2], there is an interest developing CR mimetics with the goal of extending the lifespan in humans. Energy deficits induced by CR activates AMP-activated protein kinase (AMPK) [3], which suppresses the mammalian target of rapamycin (mTOR) which is upregulated in age-related diseases such as diabetes, obesity, and cancers [4]. Metformin, an AMPK activator, and rapamycin, an inhibitor of the raptor composed of mTOR complex 1, extends the lifespan of normal mice [5,6]. We have recently reported that synthetic daumone, a pheromone secreted by Caenorhabditis elegans in response to food deficiency, improves survival in mice [7]. Considering that daumone responds to food deprivation during the dauer stage, daumone would be proposed as a CR mimetic [8].
Aging is accompanied by the continuous status of the low grade chronic inflammation [9]. Inflammation in aging is not only the result of age-related diseases but the cause of them [10]. One of the defense mechanisms against external stress is the immune system. However, continuous overstimulation of the immune system over time result in chronic inflammation, which leads to aging [9]. Studies have found that proinflammatory cytokines are increased in the plasma of aged human and experimental animals [11]. Healthy centenarians consistently show low levels of inflammatory cytokines [12]. During aging, proinflammatory molecules are up-regulated in the liver [13,14,15], and the number and phagocytic activity of Kupffer cells, which are liver macrophages, increase with aging [16]. In our previous study, daumone given at doses that extend the lifespan attenuates age-related liver injury, including inflammation and insulin resistance, in aged mice [7].
Interestingly, short-term CR largely reproduced the effects of long-term CR on the expression of inflammatory response genes in aged mice [17]. Hepatic gene expression changes associated with inflammation are down-regulated in aged mice on short-term CR for 4 weeks. These effects of short-term CR are similar those of long-term CR for 26 months. It has also been shown that short-term CR had anti-inflammatory effects on the kidneys of aged rats [18]. Activated nuclear factor-κB (NF-κB) signaling examined by binding activity and nuclear translocation in 24-month-old rat kidneys were attenuated after 10 days of CR. These data suggest that many of anti-inflammatory effects of CR are established rapidly. The present study, thus, examined the effect of late onset short-term treatment with daumone on hepatic inflammation in aged mice. We also determined the dose-response effect of daumone on tumor necrosis factor-α (TNF-α)-induced NF-κB activation in HepG2 cells.
METHODS
All chemicals were obtained from Sigma-Aldrich Co. (St. Louis, MO, USA), and tissue culture plates from NUNC (Roskilde, Denmark) and Becton Dickinson Labware (Lincoln Park, NJ, USA), unless otherwise stated. Daumone was synthesized according to the previous report [19].
Experimental animals and daumone treatment
All animal experiments were conducted according to the guidelines of the Institutional Animal Care and Use Committee (IACUC) of Ewha Womans University (2010-27-1). Six-month-old C57BL/6J male mice were purchased from Charles River Laboratory (Shizuoka, Japan), housed in a room maintained at 22±2℃, exposed to a 12-hour dark/12-hour light cycle, and fed a standard chow diet (PMI® Nutrition International, LLC Certified Rodent LABDIET® 5053, Purina Mills, Richmond, IN, USA) and autoclaved water ad libitum. Mice were maintained until the age of 24 months (old mice). Old mice were randomly divided into three groups; two groups received daumone in drinking water (2 mg/kg/day or 20 mg/kg/day) and the third received the same volume of untreated drinking water by oral gavage for 5 weeks. When daumone administration started, 9-week-old control mice (young mice) were purchased from Charles River Laboratory. Young mice were randomly divided into two groups; one received daumone (20 mg/kg/day) in their drinking water and the other received untreated drinking water. Each group of old mice contained 10 animals. Mice in the fed state were anesthetized with Avertin (intraperitoneal injection, i.p., 0.3 g/kg), and blood was collected. Livers were collected and immediately stored at -70℃.
Cell culture
HepG2 cells, human hepatoma cells, were purchased from American Type Culture Collection (Manassas, VA, USA), and maintained in Dulbecco's Modified Eagle's medium (DMEM, Invitrogen, Carlsbad, CA, USA) supplemented with 10% fetal bovine serum (FBS, Invitrogen), 100 U/mL penicillin (Invitrogen), 100 µg/mL streptomycin (Invitrogen), and 44 mM NaHCO3 at 37℃ in humidified 5% CO2 in air. Growth arrested and synchronized cells were treated with 5 ng/mL TNF-α (R&D Systems, Minneapolis, MN, USA). The cells were pretreated with daumone 6 hours before the addition of TNF-α.
Measurements of blood parameters
The blood HbA1c level was measured using the DCA2000 HbA1c reagent kit (SIEMENS Healthcare Diagnostics, Inc., Tarrytown, NY, USA). The blood was centrifuged at 900 g for 15 minutes at 4℃ and the supernatant was collected. The levels of plasma alanine transaminase (ALT) free fatty acids (FFA), total cholesterol, and triglycerides were measured using an EnzyChrom™ colorimetric assay kit (BioAssay Systems, Hayward, CA, USA).
Immunohistochemical analysis
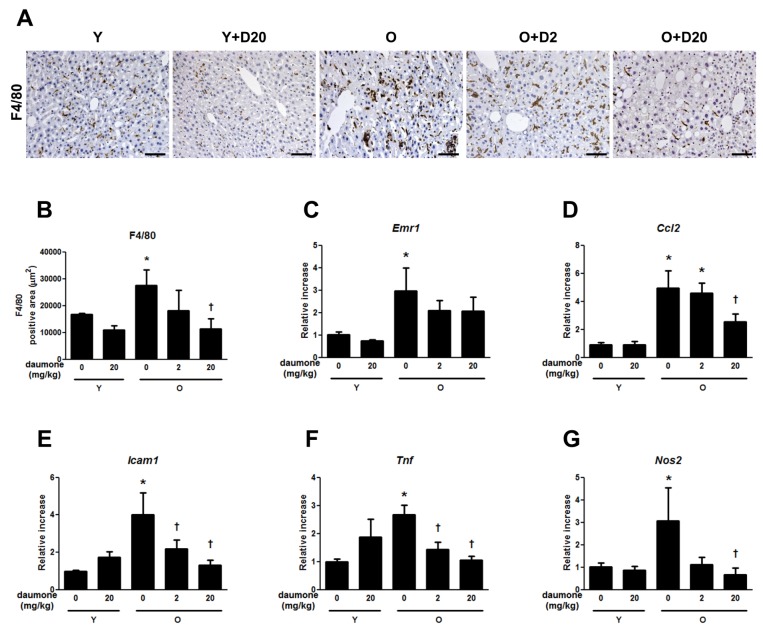
Livers were fixed in 4% formalin, dehydrated, and embedded in paraffin for sectioning. Immunohistochemistry was performed with immunoperoxidase procedures and a commercially available kit (Dakocytomation, Glostrup, Denmark). Briefly, the sections were deparaffinized, and the endogenous peroxidase activity was quenched with Dako peroxidase solution. Then, the sections were incubated with Dako serum-free blocking solution prior to incubation with anti-F4/80 (1:200 dilution, Santa Cruz Biotechnology, Santa Cruz, CA, USA) antibody overnight at 4℃. The sections were subsequently incubated with the rat ABC staining kit (Santa Cruz Biotechnology). After washing with PBS, the sections were exposed to 3,3'-diaminobenzidine. Digital images were captured on a Zeiss microscope equipped with an Axio Cam HRC digital camera (Carl Zeiss, Thornwood, NY, USA), and the positive staining area in 10 fields (100× magnification) per animal was analyzed with Image-Pro Plus 4.5.1. (Media Cybernetics, Silver Springs, MD, USA).
Real-time quantitative reverse transcription PCR (qRT-PCR)
A standard real-time qRT-PCR was performed. Total RNA was extracted from tissues using TRIzol (Invitrogen, Carlsbad, CA, USA). The mRNA levels were assessed by real-time qRT-PCR using the SYBR Green PCR Master Mix kit (Applied Biosystems, Foster City, CA, USA) with an ABI 7300 Real-time qRT-PCR thermal cycler (Applied Biosystems) as previously described [7]. The relative levels of the test genes and the internal control 18S ribosomal RNA (rRNA) were determined using a standard curve produced with the Applied Biosystems software. The primer sequences are shown in Table 1.
Western blot analysis
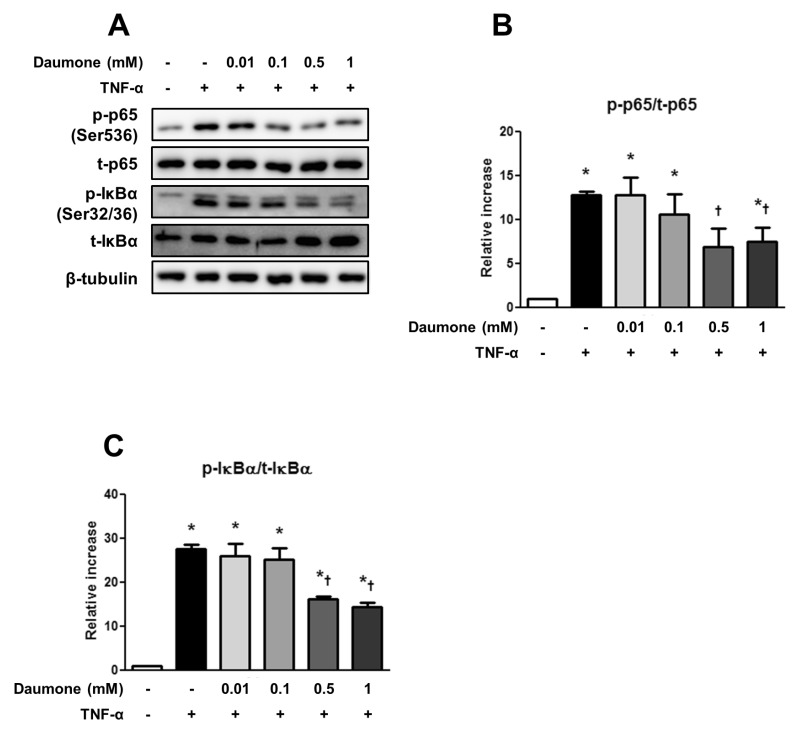
The protein levels in HepG2 cell homogenates were measured using a standard Western blotting protocol as previously described [7]. The protein concentrations of the samples were determined with the Bradford assay (Bio-Rad Laboratories, Hercules, CA, USA). Equal amounts of protein from each sample were mixed with loading buffer and separated on SDS-PAGE gels by electrophoresis. The proteins were then transferred onto a polyvinylidene fluoride membrane (GE Healthcare BioSciences, Piscataway, NJ, USA), and the membranes were incubated overnight at 4℃ with a 1:2,000 dilution of antibodies to p-NF-κB-p65 (Ser536, Cell Signaling Technology, Danvers, MA, USA), t-NF-κB (p65, Cell Signaling Technology), p-inhibitor of NF-κBα (p-IκBα, Ser32/36, Cell Signaling Technology), and t-IκBα (Santa Cruz Biotechnology). Next, the membranes were incubated with peroxidase-conjugated secondary antibodies, and the signals were visualized with an enhanced chemiluminescence system detection reagent (GE Healthcare Bio-Sciences). Positive immunoreactive bands were quantified with a densitometer (LAS-3000, FUJIFILM, Tokyo, Japan) and normalized to the levels of total protein and β-tubulin (Santa Cruz Biotechnology).
Statistical analysis
All results are expressed as the mean±standard error (SE). Analysis of variance (ANOVA) was used to assess differences between multiple groups. If the F statistic was significant, the mean values of each group were compared with the Fisher's least significant difference method. A p value<0.05 was considered statistically significant.
RESULTS
Characteristics of experimental animals
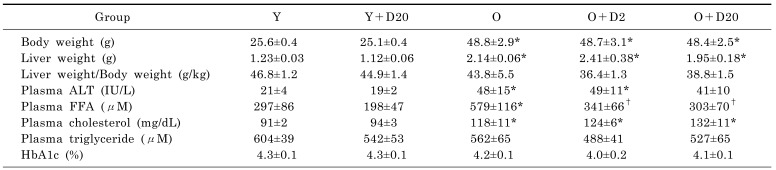
Old mice received daumone for 5 weeks from the age of 24 months by oral gavage. Twenty five-month-old mice showed significantly increased body weight and liver weight compared to young mice (Table 2). The plasma ALT of old mice was twice that of young mice. Plasma lipid such as FFA and total cholesterol were significantly increased in old mice compared to young mice. Daumone administration at doses of 2 and 20 mg/kg/day effectively decreased age-associated FFA but did not affect liver weight and plasma ALT. No significant differences were observed in plasma triglycerides and HbA1c between old and young mice.
Reduced hepatic macrophage infiltration and inflammation in the daumone-treated group
Macrophage infiltration established by F4/80 staining showed that hepatic macrophage infiltration was accelerated in old mice compared to young mice, but this was effectively reduced by the daily administration of 20 mg/kg of daumone (Fig. 1A and B). The mRNA expression of F4/80 (Emr1) was significantly increased in old mice, which was unaffected by daumone treatment for 5 weeks (Fig. 1C).
Inflammatory cytokines, including Ccl2, Icam1, Tnf, and Nos2 mRNA expression were significantly increased in the aged liver (Fig. 1D-G). Icam1 and Tnf mRNA expressions were effectively reduced by daumone administration of both 2 and 20 mg/kg/day. Ccl2 and Nos2 mRNA expressions were effectively reduced by the administration of 20 mg/kg/day of daumone.
Decreased TNF-α-induced IκBα phosphorylation in HepG2 cells treated with daumone
To assess the mechanism of the anti-inflammatory effects of daumone, the levels of TNF-α-induced NF-κB and IκBα phosphorylation in HepG2 cells were measured. TNF-α significantly increased NF-κB (Fig. 2A and B) and IκBα (Fig. 2A and C) phosphorylation, both of which were significantly suppressed by 0.5 and 1 mM daumone treatment.
DISCUSSION
The present study clearly shows that short-term treatment of daumone for 5 weeks starting in 24-month-old mice effectively attenuated age-associated hepatic inflammation in a dose-dependent manner. Cell culture study suggests inhibition of NF-κB activation as a possible underlying mechanism for the anti-inflammatory effect of daumone.
The efficacy of short-term treatment of daumone on aged mice liver was smaller than but similar with the results from late onset long-term treatment of daumone which reduced the increased histological macrophage infiltration, gene expression of Ccl2, Icam1, Tnf, and Nos2, and the risk of death and hepatic inflammation in aged mice [7]. Short-term treatment of daumone did not attenuate Emr1 (F4/80) gene expression in the liver of aged mice, unlike that of long-term daumone treatment. It will advance our understanding on molecular mechanism of daumone to examine systemic as well as hepatic inflammatory gene expression signature. The anti-inflammatory effects of daily treatment with 20 mg/kg of daumone were generally more effective than 2 mg/kg/day. Since daumone treatment up to 20 mg/kg/day for 5 weeks did not show apparent toxicity or changes in food intake, body weight change, liver function (Table 1), and behavior (data not shown), additional studies are needed to investigate whether long-term treatment with 20 mg/kg/day of daumone could more effectively improve survival and hepatic inflammation than 2 mg/kg/day [7].
Our results are consistent with previous reports demonstrating anti-inflammatory effect of short-term CR. Although it is limited, late onset CR has been suggested as a potential application to intervene and reverse age-associated organ damage [17,20,21,22]. Even a short duration of CR for 10 days has anti-inflammatory effects on 24-month-old rat kidneys [18], and CR for 4 weeks as well as lifelong CR reversed the majority of age-associated gene expression in 34-month-old mice livers [17].
Hepatic injury estimated by increased ALT was apparent in 25-month-old mice, and was not altered by short-term daumone treatment, which differs from the results of long-term daumone treatment [7]. Three months of CR reduced senescence associated with β-galactosidase (SA-β-gal) activity and improved telomere maintenance in 14-month-old mice livers [21], suggesting that the anti-inflammatory effect may be the early therapeutic benefit of CR mimetics or anti-aging agents. Experiments like these should be utilized to quickly identify effective CR mimetics or anti-aging agents.
The anti-inflammatory effects of daumone may result from inhibition of NF-κB. Lifelong CR attenuates nuclear NF-κB and cytosolic IκBα phosphorylation in 24-month-old mice livers [15]. Similarly, long-term treatment of daumone reduces cytosolic IκBα phosphorylation in aged mice livers [7]. In the present study, increased IκBα and NF-κ B-p65 phosphorylation in TNF-α-treated HepG2 cells were concentration-dependently reduced by daumone treatment. It remains to be determined whether daumone affects macrophage NF-κB activation. NF-κB activation in macrophages leads to upregulate inflammatory cytokines including TNFα [23,24]. However, inhibition of macrophage NF-κB through macrophage-restricted deletion of IκB kinase 2 increases atherosclerosis [25].
The present data demonstrated for the first time that late onset short-term treatment of daumone, a possible CR mimetic, has protective effects on liver inflammation through the suppression of NF-κB activation, and suggests that daumone may become a lead compound in the treatment of age and age-related diseases.

 XML Download
XML Download