

 PDF
PDF ePub
ePub Citation
Citation Print
Print


ABBREVIATIONS
COX
cyclooxygenase
ERK1/2
extracellular signal-regulated kinase 1/2
iNOS
inducible nitric oxide synthase
IL1β
interleukin 1β
JNK
c-Jun N-terminal kinase
LPS
lipopolysaccharide
MAPK
mitogen-activated protein kinase
NF-κB
nuclear factor-kappa B
NO
nitric oxide
RANKL
receptor activator of nuclear factor kappa-B ligand
TNF-α
tumor necrosis factor-α
INTRODUCTION
Silymarin is an extract isolated from the fruit and seeds of milk thistle Silybum marianum [1]. Previous studies showed that silymarin protected against hepatotoxicity caused by a variety of toxins and chemicals including microcystin, ochratoxin, ethanol, phenylhydrazine, and acetaminophen [2,3,4,5]. High-performance liquid chromatography (HPLC) and capillary zone electrophoresis analyses have identified the main components of silymarin as silibinin, isosilybin, silydianin, and silychristin [6]. Silibinin, a major polyphenolic flavonoid component of silymarin, has shown a number of additional biological activities such as anti-inflammatory and anti-carcinogenic effects [7,8,9,10].
Although the mechanisms underlying the effects of silymarin are largely unknown, possible targets have been suggested. Silibinin and related compounds are direct inhibitors of hepatitis C virus RNA-dependent RNA polymerase, suggesting potential for the treatment of hepatitis C [11]. Isosilybin A was shown to act as a partial agonist of the peroxisome proliferator-activated receptor type gamma, a current therapeutic target in type 2 diabetes and metabolic syndrome [12]. Silymarin has antioxidant activity, scavenging free radicals and modulating antioxidant and inflammatory enzymes [13,14]. It has also been reported that mitogen-activated protein kinases (MAPKs) are potential targets of silymarin and silibinin [15,16]. Silymarin inhibits MAPKs and induces apoptosis in established skin tumors in SENCAR mice [15]. Silibinin prevents the activation of MAPKs and nuclear factor-kappa B (NF-κB) in osteoclast precursors in response to RANKL [17].
LPS stimulation of macrophages induces morphological changes, including an increase in cell size, production of lamellipodia and filopodia, and alterations in cell adhesion, migration, and phagocytosis [18]. Lamellipodia are thin sheets of cell edges containing large amounts of actin filaments and filopodia are actin-containing spikes [19]. Cytoskeletal and adhesion dynamics which mediate migration are regulated by Rho GTPases including RhoA, the Rac subfamily, and Cdc42 [20]. NF-κB also regulates the actin cytoskeleton via integrin-mediated signal transduction, and consequently induces morphological changes in these cells, including lamellipodia formation [21].
Due to the important role of macrophages in inflammation, suppression of their LPS-induced morphological changes and production of inflammatory mediators is an effective way to prevent a variety of inflammatory disorders.
Go to : 
METHODS
Materials
Silymarin, BAY-11-7085, and LPS from Salmonella thyposa were purchased from Sigma (St. Louis, MO). The anti-iNOS antibody was purchased from Upstate Biotechnology (Lake Placid, NY, USA) and antibodies against phospho-p44/42, p44/42, phospho-p38, p38, phospho-JNK, and JNK were purchased from Cell Signaling Technology, Inc. (Beverly, MA, USA).
Cell culture
RAW 264.7 cells were purchased from the American Type Culture Collection (Bethesda, MD, USA). Cells were grown in DMEM supplemented with 10% fetal bovine serum, 2 mM L-glutamine, 100 U/ml penicillin, and 100 mg/ml streptomycin. Cells were cultured in the presence of 5% CO2 at 37ℰ.
Morphological analysis
The RAW 264.7 cells (5×105 cells/ml) were cultured in the presence of silymarin and LPS for 3, 6, or 12 h on cover slides in 12-well plates. Morphological changes were analyzed by light microscopy and Giemsa staining. Giemsa's solution is a mixture of methylene blue, eosin, and azure B. It specifically attaches to the phosphate groups of DNA, where there are high amounts of adenine-thymine bonding.
Cell adhesion assay
The cell adhesion assay was performed as described previously [22], with minor modifications. RAW264.7 cells were treated with silymarin (50 μg/ml) in the presence of LPS (200 ng/ml) for 18 h. Cells were collected and re-plated at a density of 1×105 cells/ml. After 30 min, unattached cells were removed by washing with phosphate-buffered saline (PBS) 3 times. Cell adhesion was calculated by counting the attached cells and expressing this number as a percentage of the total cells.
Nitrite determination
RAW264.7 cells were treated with the indicated concentrations of silymarin in the presence of LPS (200 ng/ml) for 18 h. Culture supernatants were collected, and the accumulation of nitrite in culture supernatants was measured as an indicator of NO production in the medium, as previously described [23,24,25].
Western immunoblot analysis
Whole cell lysates were separated by 10% SDS-PAGE and then electrotransferred to nitrocellulose membranes (Amersham International, Buckinghamshire, UK). The membranes were then preincubated for 1 h at room temperature in Tris-buffered saline (TBS), pH 7.6, containing 0.05% Tween-20 and 3% bovine serum albumin, followed by incubation with primary antibody. Immunoreactive bands were detected by incubation with conjugates of anti-rabbit IgG with horseradish peroxidase and enhanced chemiluminescence reagent (Amersham).
RT-PCR
Total RNA was isolated with TRI Reagent (Molecular Research Center, Cincinnati, OH, USA). Forward and reverse primer sequences were as follows: iNOS: 5'-CTG CAG CAC TTG GAT CAG GAA CCT G-3', 5'-GGG AGT AGC CTG TGT GCA CCT GGA A-3'; and β-actin: 5'-TGG AAT CCT GTG GCA TCC ATG AAA C-3', 5'-TAA AAC GCA GCT CAG TAA CAG TCC G-3'. Equal amounts of RNA were reverse-transcribed into cDNA with oligo (dT) 15 primers. PCR was performed with cDNA and each primer. Samples were heated to 94ℰ for 5 min and then cycled 30 times at 94ℰ for 1 min, 55ℰ for 1.5 min, and 94ℰ for 1 min, after which an additional extension step at 72ℰ for 5 min was conducted. PCR products were separated by 8% SDS-PAGE, followed by staining with ethidium bromide. The iNOS and β-actin primers produced amplified products of 311 and 349 bps, respectively.
Luciferase reporter gene assay
RAW264.7 cells were transfected with the pNF-κB-Luc plasmid. Cells were incubated for 24 h and then treated with silymarin (50 μg/ml) in the presence of LPS for 18 h. Cell extracts were prepared and luciferase activity was analyzed to enable determination of NF-κB transactivation activity.
Immunofluorescence staining
RAW264.7 cells were treated with silymarin (50 μg/ml) in the presence of LPS on a cover slide in 12-well plates. Cells were rinsed 3 times with PBS, fixed with 4% paraformaldehyde for 10 min at room temperature, and rinsed again. Cells were then blocked with 1% bovine serum albumin, followed by the addition of the primary antibody. After extensive washing with TBS, fluorescein isothiocyanateconjugated IgG was added. Following incubation, the slides were rinsed, mounted, and viewed at 488 nm on a confocal microscope (FV300, Olympus, Japan).
Statistical analysis
The mean±standard deviation (SD) was determined for each treatment group in a given experiment. To determine whether significant differences were present, treatment groups were compared to the respective vehicle controls using Student's two-tailed t test.
Go to :
RESULTS
Inhibition of macrophage activation by silymarin in LPS-stimulated RAW264.7 cells
To investigate the effects of silymarin on macrophage activation, RAW 264.7 cells were analyzed by light microscopy, because LPS-induced morphological change is a hallmark of macrophage activation. The RAW 264.7 cells (5×105 cells/ml) were cultured in the presence of silymarin and LPS for 3, 6, or 12 h on cover slides in 12-well plates. LPS stimulation for 6 h and 12 h induced lamellipodia extension and spreading of cells (Fig. 1A). However, unstimulated cells did not show morphological changes at any time point. Silymarin treatment of RAW 264.7 cells prevented LPS-induced morphological changes at 6 h and 12 h in a dose-dependent manner (Fig. 1A). We counted total cells and fully spread cells and calculated fully spread cell percentage. Counting of the fully spread cells further showed that silymarin inhibited LPS-induced morphological changes in this macrophage cell line (Fig. 1B). These data suggested that silymarin inhibited activation of LPS-stimulated RAW 264.7 cells.
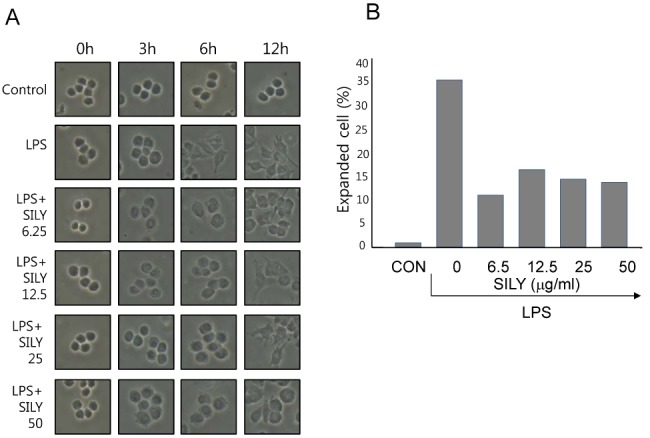 | Fig. 1Inhibition of macrophage activation by silymarin in LPS-stimulated macrophages. RAW264.7 cells were treated with the indicated concentrations of silymarin in the presence of LPS (200 ng/ml) for 3 h, 6 h, or 12 h on cover slides in 12-well plates. Cells were then subjected to microscopic analysis. (A) Representative photographs. (B) In 12 h LPS treatment samples, fully expanded cells were counted and expressed as a percentage of the total number of cells.
|
Inhibition of morphological change and adhesion activity by silymarin in LPS-stimulated macrophages
Giemsa staining of RAW264.7 cells treated with LPS for 6 h showed that lamellipodia were extended from 35 sides of the cells, and that silymarin inhibited lamellipodia formation (Fig. 2A). To further analyze whether the inhibition of lamellipodia formation by silymarin was related to cell adhesion activity, we investigated the effects of silymarin on cell attachment to culture plates. LPS stimulation significantly increased RAW264.7 adhesion, whereas silymarin inhibited this activity (Fig. 2B). Microscopic analysis showed that LPS-stimulated macrophages were attached after washing, while control and silymarin treated macrophages were washed off the plate (Fig. 2C). We further analyzed the effects of silymarin on morphological changes using immunofluorescence staining of F-actin. Silymarin inhibited lammellipodia and filopodia formation induced by LPS (Fig. 3). These results demonstrated that silymarin inhibited both the morphological changes and functional activation of mouse RAW264.7 macrophages.
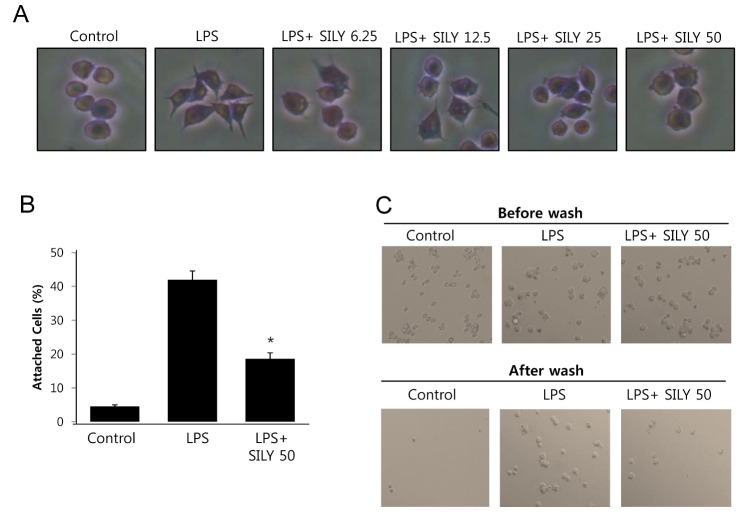 | Fig. 2Inhibition of morphological change and adhesion activity by silymarin in LPS-stimulated macrophages. (A) RAW264.7 cells were treated with the indicated concentrations of silymarin in the presence of LPS (200 ng/ml) for 3 h, 6 h, or 18 h on cover slides in 12-well plates. Cells were then Giemsa-stained and analyzed for morphological change. (B) RAW264.7 cells were treated with silymarin (50 μg/ml) in the presence of LPS for 24 h. Cells were harvested, washed, plated in 6-well plates (5×105/ml) for 30 min. Cells were then washed with PBS and analyzed for cell attachment using microscopy. Attached cells were counted after washing. (C) Representative photographs of attached cells before and after washing. Each column shows the mean±SD of triplicate determinations. *p<0.05 compared to the control group, as determined by Student's two-tailed t test.
|
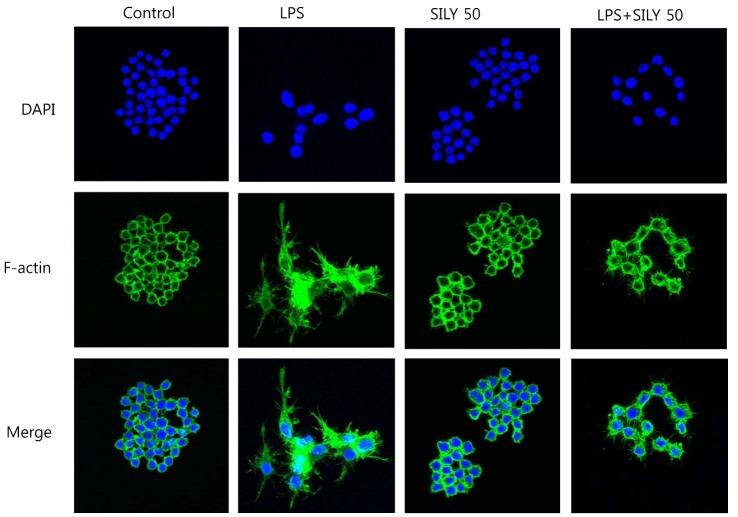 | Fig. 3Inhibition of the lamelipodia and filopodia formation by silymarin in LPS-stimulated macrophages. RAW 264.7 cells were treated with the silymarin (50 μg/ml) in the presence of LPS for 18 h on cover slides in 12-well plates. Cells were then subjected to immunofluorescence staining of F-actin and DAPI staining.
|
Inhibition of NF-κB activation and p65 nuclear translocation by silymarin in LPS-stimulated macrophages
The effect of silymarin on NF-κB, which binds the iNOS gene promoter [26], was evaluated using a luciferase reporter assay. Treatment of RAW264.7 cells with LPS induced a marked increase in NF-κB-induced luciferase activity, whereas the induction of NF-κB activity was inhibited by silymarin (Fig. 4A). Macrophage stimulation induces nuclear translocation of p65, one of the major components of NF-κB, which binds to the promoter and induces transcription. We therefore analyzed the effect of silymarin on the nuclear translocation of p65 using immunofluorescence staining. When RAW264.7 cells were stimulated by LPS for 2 h, p65 levels increased in the nucleus, whereas p65 was mainly localized in the cytosol of unstimulated control cells (Fig. 4B). This LPS-induced nuclear translocation of p65 was inhibited by silymarin. Western immunoblot analysis further showed that silymarin reduced p65 in the nucleus (Fig. 4C). These results indicated that silymarin reduced the nuclear translocation and transactivation activity of NF-κB, which is important in LPS-mediated regulation of inflammatory gene expression in macrophages.
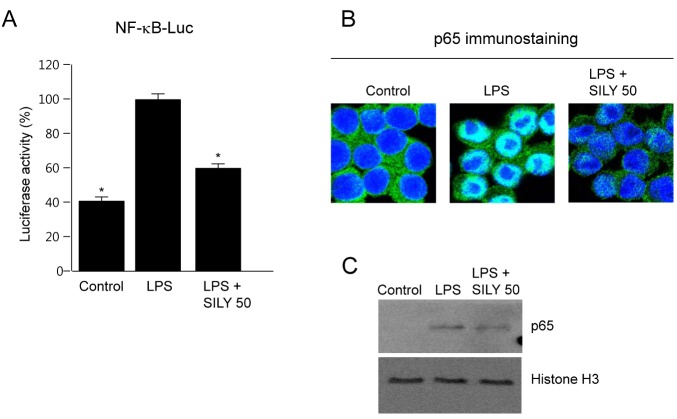 | Fig. 4Inhibition of nuclear factor-kappa B (NF-κB) activation and p65 nuclear translocation by silymarin in LPS-stimulated macrophages. (A) RAW264.7 cells were transfected with pNF-κB-Luc plasmid. Cells were incubated for 24 h and then treated with silymarin (50 μg/ml) in the presence of LPS for 18 h. Cell extracts were prepared and luciferase activity was analyzed to indicate NF-κB transactivation activity. (B) RAW264.7 cells were treated with silymarin (50 ±g/ml) in the presence of LPS for 2 h on cover slides in 12-well plates. Cells were then subjected to immunofluorescence staining of p65 and DAPI staining. (C) RAW264.7 cells were treated with silymarin (50 μg/ml) in the presence of LPS for 2 h. Nuclear extracts were then prepared and subjected to Western immunoblot analysis using anti-p65 antibody. Each column shows the mean±SD of triplicate determinations. *p<0.05 compared to the control group, as determined by Student's two-tailed t test.
|
Inhibition of NF-κB blocks morphological changes in LPS-stimulated macrophages
To further confirm the involvement of NF-κB activation in the morphological changes, we analyzed the effects of a pharmacological inhibitor of NF-κB on macrophage activation using microscopy and a nitrite generation assay. BAY-11-7085, an inhibitor of IκBα phosphorylation, inhibited LPS-induced nuclear translocation of p65 (Fig. 5A). Treatment of RAW264.7 cells with BAY-11-7085 significantly inhibited LPS-induced nitrite production, one of indicator of macrophage activation [27], and morphological changes in a dose dependent manner (Fig. 5B, 5C). These results demonstrated that NF-κB was important for both the morphological and functional activation of macrophages.
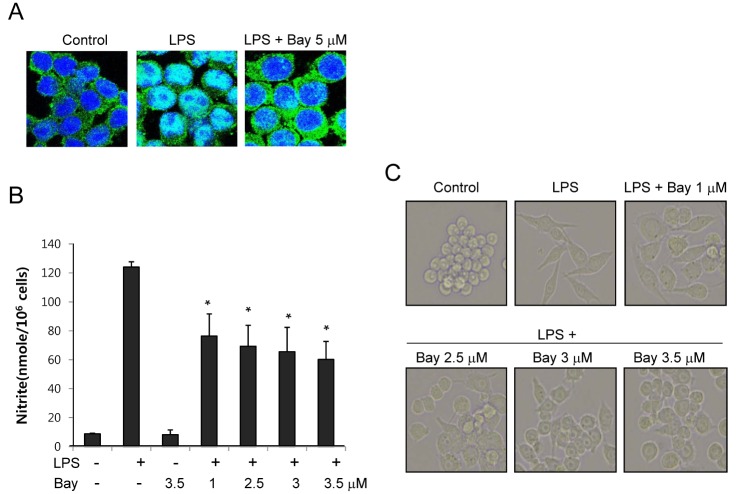 | Fig. 5Effects of nuclear factor-kappa B (NF-κB) inhibitor on macrophage activation induced by LPS. (A) RAW264.7 cells were treated with BAY-11-7085 (5 μM) in the presence of LPS for 2 h on cover slides in 12-well plates. Cells were then subjected to immunofluorescence staining of p65 and DAPI staining. (B) RAW 264.7 cells were treated with the indicated concentrations of BAY-11-7085 in the presence of LPS for 24 h. Supernatants were subsequently isolated and analyzed for nitrite. Each column shows the mean±SD of triplicate determinations. *p<0.05 compared to the control group, as determined by Student's two-tailed t test. (C) RAW264.7 cells were treated with the indicated concentrations of BAY-11-7085 in the presence of LPS (200 ng/ml) for 6 h on cover slides in 12-well plates. Cells were then subjected to microscopic analysis.
|
Inhibition of MAPKs phosphorylation by silymarin in LPS-stimulated RAW264.7 cells
Since MAPKs are important for NO generation in LPS-stimulated RAW264.7 and are potential targets of silymarin, we further determined the role of MAPKs in the inhibition of NO by silymarin. RAW264.7 cells were pretreated with silymarin for 1 h and then incubated for 30 min in the presence of LPS. MAPK phosphorylation was analyzed by western blot. The phosphorylation of ERK1/2 was strongly increased by LPS treatment, whereas basal ERK1/2 phosphorylation was undetectable (Fig. 6A). When cells were treated with silymarin in the presence of LPS, ERK1/2 phosphorylation was decreased in a dose-dependent manner (Fig. 6B). The phosphorylation of c-Jun N-terminal kinase (JNK) was also increased by LPS and decreased by silymarin (Fig. 6C). Western blot of p38 showed modest basal phosphorylation, slight induction by LPS, and inhibition by silymarin (Fig. 6C).
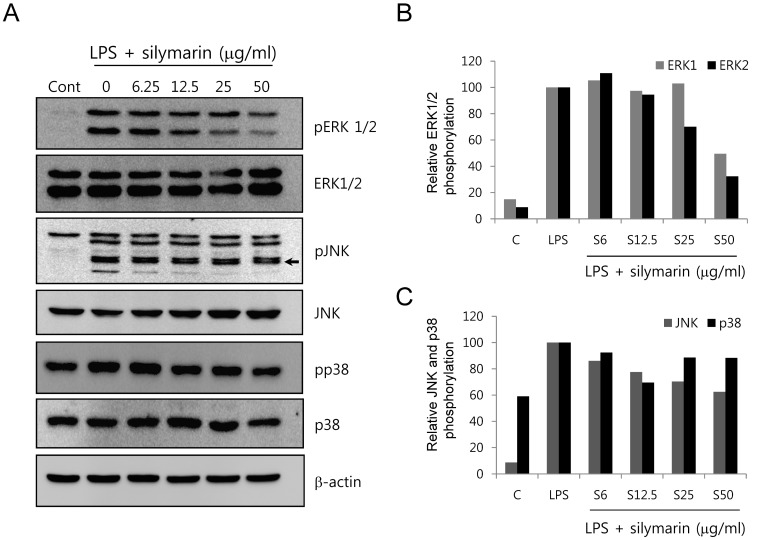 | Fig. 6Inhibition of mitogen-activated protein kinase (MAPK) phosphorylation by silymarin in LPS-stimulated RAW264.7 cells. (A) RAW264.7 cells were pretreated with silymarin. RAW264.7 cells were pretreated with silymarin for 1 h and incubated for 30 min in the presence of LPS. The phosphorylation of extracellular signal-regulated kinase 1/2 (ERK1/2), p38, and phospho-c-Jun N-terminal kinase (pJNK) was analyzed by Western blot. The relative band densities of ERK1/2 (B) and p38 and pJNK (C) were analyzed using the Image J program.
|
Go to :
DISCUSSION
The present study showed that silymarin, a polyphenolic flavonoid isolated from milk thistle (Silybum marianum), was a potent anti-inflammatory agent. Silymarin inhibited morphological changes, such as lamellipodia and filopodia formation, in LPS-stimulated RAW264.7 cells. Lamellipodia and filopodia are thin sheets of cell edges containing large amounts of actin filaments and actin-containing spikes, respectively [19]. These morphological changes are the hallmarks of the macrophage inflammatory response, because formation of lamellipodia and filopodia is important for attachment to the extracellular matrix, which is required for cell adhesion and migration to inflammatory sites. The anti-inflammatory effect of silymarin was further supported by its inhibition of NO production, an important mediator of inflammatory responses, including phagocytosis [8]. The inhibition of NO production was related to the suppression of iNOS gene expression. Macrophages play an important role in inflammation and in the pathogenesis of autoimmune diseases, and the inhibition of morphological and functional changes of macrophages by silymarin therefore indicated its potential to act as an anti-inflammatory agent. Our data, and those of others, have demonstrated that silymarin protected pancreatic beta cells from destruction induced by proinflammatory cytokines [25,28]. Proinflammatory cytokines are known to induce the expression of iNOS mRNA and the production of NO, resulting in beta cell death [29,30] An in vivo study using a rat model further showed that silymarin increased insulin gene expression and beta cell proliferation [31].
One of the most important findings of the present study was the identification of the involvement of NF-κB in the macrophage morphological changes induced by LPS. We showed that silymarin reduced nuclear translocation of p65, a component of NF-κB. In unstimulated cells, a heterodimer of p65 and p50 (another NF-κB component) is bound to IκB, an inhibitor of NF-κB. NF-κB is released by LPS-induced phosphorylation of IκB and can then translocate to the nucleus. Because p65 has transactivation activity, the nuclear translocation of p65 into the nucleus is a key process in the activation of NF-κB. Further, silymarin inhibited NF-κB-dependent luciferase reporter gene activity. Our inhibitor study suggested that there was a relationship between NF-κB and macrophage morphological changes. BAY-11-7085, an inhibitor of IκBαphosphorylation, inhibited the nuclear translocation of p65 and morphological changes in LPS-stimulated macrophages. The role of NF-κB in morphological changes is further supported by previous reports [21]. NF-κB regulated the actin cytoskeleton and induced morphological changes, including lamellipodia formation [21]. NF-κB also plays an important role in gene expression of inflammatory mediators including iNOS, COX-2, and cytokines [32]. Our data also showed that silymarin and an NF-κB inhibitor prevented NO production. The possible targets of NF-κB in mediating morphological changes are cell adhesion molecules, such as VCAM-1, ICAM-1, and E-selectin [33,34,35]. Silymarin has been reported to inhibit monocyte adhesion to endothelial cells and the gene expression of cell adhesion molecules in endothelial cells when monocytes were activated [36].
To investigate the mechanism by which silymarin inhibited macrophage activation, we analyzed the effects of silymarin on MAPK pathways. Macrophage stimulation by external stimuli, including LPS, induces the phosphorylation of MAPK family members including ERK1/2, p38, and JNK [37]. These are serine threonine kinases that are located in the cytoplasm until activated by dual phosphorylation of their Thr and Tyr residues at Thr-Glu-Tyr, Thr-Gly-Tyr, or Thr-Pro-Tyr, respectively [38]. We showed that silymarin inhibited ERK1/2, JNK, and p38 phosphorylation. ERK2 phosphorylation was most strongly inhibited by silymarin (Fig. 6). Previous studies showed that MAPKs were potential targets of silymarin and silibinin [15,17]. Silymarin inhibited growth and caused regression of skin tumors in SENCAR mice through the inhibition of MAPKs [15]. Silibinin prevented RANKL-induced activation of ERK, p38, JNK, and NF-κB in osteoclast precursor cells [17]. Silibinin inhibited TPA-induced MMP-9 expression through the Raf/MEK/ERK pathway in thyroid and breast cancer cells [39,40]. Silibinin also prevented TNF-αinduced MMP-9 expression in gastric cancer cells through inhibition of the MAPK pathway [16]. ERK activity is required for iNOS gene expression in insulin-producing INS-1E cells and RAW 264.7 cells [41,42]. Although we and many other researchers have shown that silymarin inhibited MAPK phosphorylation, the direct targets of silymarin need to be identified. The upstream kinases, including MAP/ERK kinase (MEK)1/2, MAP kinase kinase (MKK)3/6, and MKK4/7, should be tested first as candidate silymarin targets, although we could not exclude the possibility that MAP3Ks are silymarin targets.
The present study demonstrated that silymarin inhibited MAPK pathways and NF-κB in LPS-stimulated macrophages. Although MAPKs and NF-κB play pivotal roles in macrophage activation, including lamellipodia and filopodia formation, the exact relationships between these pathways and the morphological changes are largely unknown. However, some proteins are known to be involved in the control of lamellipodia and filopodia formation. The complex of actin-related proteins 2/3 (Arp2/3) is a major initiator of new actin filaments in cells and plays an essential role in the assembly of actin networks in lamellipodia. Filopodia are controlled by proteins such as fascin, diaphanous, and mena/vasodilator-stimulated phosphoprotein (VASP). Fascin is an actin bundling protein that brings together filaments in lamellipodial networks to encourage filopodial growth [43]. Its actin bundling activity is regulated by phosphorylation and by the small GTPases, Rac and Cdc42 [19,44,45]. Murine diaphanous is involved in different cytoskeletal activities, including its interaction with microtubules [46]. Mena/VASP proteins have the ability to promote the elongation of long parallel bundles of actin in filopodia [47]. All of these proteins have been implicated in lamellipodial and filopodial actin dynamics, so it seems that silymarin may control these proteins, directly or indirectly, through the regulation of MAPKs and NF-κB pathways.
Collectively, silymarin exerted potent control over pathophysiological processes involved in the inflammatory response through the inhibition of MAPKs and NF-κB pathways. Due to the critical roles of these pathways in mediating adhesion and migration of cancer cells, silymarin represents a possible chemotherapeutic agent.
Go to :
 XML Download
XML Download