

 PDF
PDF ePub
ePub Citation
Citation Print
Print


ABBREVIATIONS
Asp
aspartic acid
CNS
central nervous system
DA
dopamine
DOPAC
3,4-dihydroxyphenylacetic acid
ELF-MF
extremely low frequency magnetic field
GABA
γ-aminobutyric acid
Gln
glutamine
Glu
glutamate
Gly
glycine
HIAA
5-hydroxyindoleacetic acid
NE
norepinephrine
NO
nitric oxide
Tyr
tyrosine
VMA
vanillyl mandelic acid
5-HT
serotonin
INTRODUCTION
Since the electricity became the prevalently used source of energy, we are exposed to extremely low frequency magnetic field (ELF-MF) almost everywhere in our environment. ELF-MF is classified by their wave lengths and frequencies [1,2]. On the issue of biological effects and medical applications of ELF-MF, many articles have been published over the past three decades. Based on a recent in-depth review of the scientific investigations [3,4,5,6], it is concluded that current evidence does not confirm the existence of any health consequences from exposure to low level ELF-MF. However, it is also acknowledged that understanding of ELF-MF and its influences on health conditions needs further scientific research.
Several epidemiological studies reported an elevated cancer risk, especially for childhood leukemia and brain cancer, associated with exposure to residential and occupational ELF-MF, whereas other studies have not confirmed these results [7,8]. Investigations were also conducted on the possible involvement of the melatonin circadian secretion profile in ELF-MF effects [9]. In fact, it has been suggested that the alteration of the blood level of this hormone consequent to ELF-MF exposure could result in a decreased protection against tumor growth [10,11]. On the other hand, studies on the central nervous system (CNS) showed a depressed activity of different types of neurotransmitter system and protein content in the brain of animals exposed to ELF-MF [12,13,14,15].
The CNS is a potential site of interaction between biological systems and ELF-MF because of the electrical sensitivity of the tissues. Several studies have identified occupational exposure to ELF-MF as a potential risk factor for neurodegenerative disease [16,17]. They discussed that ELF-MF may interfere with the neuropsychological processes responsible for this short-term learning effect supported by brain synaptic plasticity. Clinical electroencephalogram (EEG) measurements during ELF-MF were reported to show desynchronization of the alpha rhythm (the dominant frequency range in the human brain, 8~12 Hz) and other abnormalities in EEG activity [18,19]. The EEG and neurotransmitters have an intimate relationship [20,21,22]. When action potentials are triggered simultaneously by neurons, giving rise to detectable EEG signals, it triggers synaptic transmission and neurotransmitters release [23]. Therefore, the alteration of EEG or neurotransmission by ELF-MF may be of interest.
In this study, we described the influence of ELF-MF on the level of various neurotransmitters of a rat brain. In addition, we investigated the concentration of NO in several brain areas of rats that were exposed to ELF-MF. Through this study we aimed to get more information in elucidating the effects of ELF-MF on neurotransmission in CNS.
Go to : 
METHODS
Animals
Male Sprague-Dawley rats (Samtako, Osan, Korea) weighing 250~350 g were used in all experiments. These rats were treated in accordance with the National Institutes of Health (NIH, Bethesda, MD) Guide for the Humane Care and Use of Laboratory Animals and Chung-Ang University (Seoul, Korea) Institutional Animal Care and Use Committee guidelines. Animals were maintained in a temperature-controlled room (25±2℃) and kept on a 12:12 lightdark cycle (lights on at 8:00 a.m. KST). Food and water were available ad libitum and the rats were acclimated for 2 weeks to these conditions before the experiment. After ELF-MF exposure, the rats were anesthetized with sodium pentobarbital (50 mg/kg, i.p.). The brain regions of interest were rapidly removed, weighed to the nearest milligram, frozen rapidly in microcentrifuge tubes, and stored at -70℃.
ELF-MF generation
The 60 Hz ELF-MF was produced by one pair of Helmholtz coils with windings embedded in an open rectangular frame (140×85×70 cm3). Each coil had 200 turns of an insulated soft copper wire with a diameter of 1.0 mm, and was connected in series to a 220 V AC power supply via a transformer. Each winding was split, allowing the current to flow in the same direction through each half of the winding (field adding). We modulated the intensity of ELF-MF using the transformer. These coils produced a 60 Hz magnetic field set to 2.0 mT root mean square (rms) amplitude using a gaussmeter (Model 410; Lake Shore Cryotronics, Westerville, OH) in the vertical direction at the center of the system. The magnetic field intensity was not uniform in the range of 1.8~2.0 mT rms, depending on the position of the magnetic induction measurement points. The measurements were taken at five points (one at the center and four at the edges of the cage location). The highest values of magnetic field induction (2.0 mT rms) were found at the center and the lowest ones (1.8 mT rms) at the edge of cage. The non-magnetic cage was placed at the center of the cage location.
Exposure Conditions
Rats were assigned into one of two groups (sham group, n=10 and each experimental group, n=10). The experimental groups were exposed to 2.0 mT, 60 Hz ELF-MF for 2 (n=10) or 5 days (n=10) in the cage. The sham control animals were placed in the same location with the coils turned off and were exposed to the local ambient geomagnetic field (BH=18, BV=22, BT=23 and BAC<2 µT, where BH is the horizontal magnetic field intensity, BV is the vertical magnetic field intensity, BT is the total magnetic field intensity, and BAC is the alternating magnetic field intensity).
Tissue preparation
Each rat was killed under anesthesia after the exposure to ELF-MF or sham condition. The brain was removed and dissected into five parts of brain (cerebellum, cortex, hippocampus, thalamus and striatum) for HPLC and NO assay. The dissected tissue were quickly frozen on dry ice, weighed and stored in 1.5 ml microtubes at -70℃.
HPLC analysis
HPLC analysis system consisted of pump (Model PU-980 Pump, Jasco, Japan), reverse phase column (Luna C18, 4.6 mm I.D. ×250 mm length, 5 µm particle size, Phenomenex, USA), ECD detector (JASCO MODEL 840-EC). For biogenic amines and their metabolites, mobile phase was 0.75 mM otanesulforic acid, 0.07 M NaH2PO4 and 0.1 mM EDTA in 12% methanol solution. The pH of the mobile phase was adjusted to 2.8 with perchloric acid and then degassed for 15 min in sonicator (Branson 3200). Mobile phase for amino acids consisted of 0.1M NaH2PO4, 0.5 mM EDTA in 25% methanol solution adjusted to pH 4.5 with 1M phosphoric acid. After decapitation, brain was rapidly removed and dissected into various regions on a cold dissecting plated as previously described. All samples were sonicated in 0.1 mol perchloric acid solution containing an internal standard. Dihydroxybenzylamine and β-aminobutyric acid were used as internal standard for biogenic amines and amino acids, respectively. The sonicated tissue was centrifuged at 13,000×g for 20 min and the supernatant (20 µl) was injected into HPLC. This system was run at voltage electrodes set at +670 mV. 10 nmol and flow rate was fixed at 1 ml/min.
Brain NOx assay
Tissue NOx contents were determined as described by Tracey et al. [24]. Brain samples were weighted and homogenized in 400 µl deionized water, and centrifuged at 20000 g, for 10 min, at 4℃. Fifty microlitres of supernatant was mixed with 20 µl of 0.31 M potassium phosphate buffer (pH 7.5), 10µl of 0.86 mM β-NADPH, 10µl of 0.11 mM FAD and 20 mu of nitrate reductase. Samples were allowed to incubate for 1 h at room temperature in the dark. Then, 5 µl of 1M ZnSO4 were added to the samples in order to precipitate the proteins. Samples were centrifuged at 20000 g, for 5 min, at 4℃ and the supernatants were removed. One hundred microlitres of Griess reagent (1:1 mixture of 1% sulphanylamide in 5% H3PO4 and 0.1% N-(-1-naphtyl) ethylenediamine) were added to 50 µl of supernatant and the mixture incubated for 10 min at room temperature. Absorbances were measured at 540 nm by a spectrophotometer and converted to NOx content by using a nitrate standard curve.
Data expression and statistical analysis
Within each groups, the mean±standard errors of each neurotransmitters were compared. Statistical significance of the differences between the groups was determined by using Student's t test or ANOVA after verifying the fitness for parametric analysis.
Go to :
RESULTS
Determination of biogenic amine neurotransmitters and their metabolites
The exposure of 2.0 mT ELF-MF for 2 (n=10) or 5 days (n=10) produced the decrease in norepinephrine (NE) level in most of brain area (Table 1). Significant differences in the NE level were found in the striatum, thalamus, cerebellum and hippocampus (p<0.01). In the same areas in which NE was affected by ELF-MF, the concentration of the metabolite of NE, VMA, was also found significantly higher than in those of sham control (p<0.01). In contrast, there was no change in NE and its metabolite in the cortex following exposure to 2.0 mT ELF-MF for 2 or 5 day.

The exposure to 2.0 mT ELF-MF caused a significant increase of serotonin (5-HT) in the striatum and thalamus (p<0.05; Table 1). The increases occurred only in the group exposed to ELF-MF for 5 days, but not for 2 days. The level of metabolite of 5-HT, HIAA, was shown to change significantly in these brain areas by ELF-MF. The significant increase of HIAA by exposure to ELF-MF for 5 days occurred in the striatum, thalamus (p<0.01) and hippocampus (p<0.05). In the thalamus, 2 day long exposure to magnetic fields also produced the increase of HIAA (p<0.01).
Dopamine (DA) was detected only in the striatum and thalamus (Table 1). In the striatum, higher level of DA was detected compared to thalamus. The exposure to 2.0 mT ELF-MF did not induce any change of this neurotransmitter and its metabolites, DOPAC and HVA, in the striatum. The exposure to ELF-MF for 5 days significantly increased the concentration of DA in the thalamus (p<0.05). Although both metabolites of DA, DOPAC and HVA, was detected in the striatum or thalamus, there was no alteration in their levels by exposure to ELF-MF.
Determination of amino acid neurotransmitters
The exposure to ELF-MF altered the level of amino acid neurotransmitters in the brain (Table 2). ELF-MF tended to increase the levels of amino acid neurotransmitters in the striatum and thalamus, while it decreased the levels in the cortex, cerebellum and hippocampus. In the thalamus, the significant increases in glutamate (Glu), glutamine (Gln), glycine (Gly), tyrosine (Tyr) and GABA were observed after exposure to ELF-MF for 5 days (p<0.05). Gln, Gly and GABA were also elevated in the striatum by ELF-MF exposure (p<0.05).
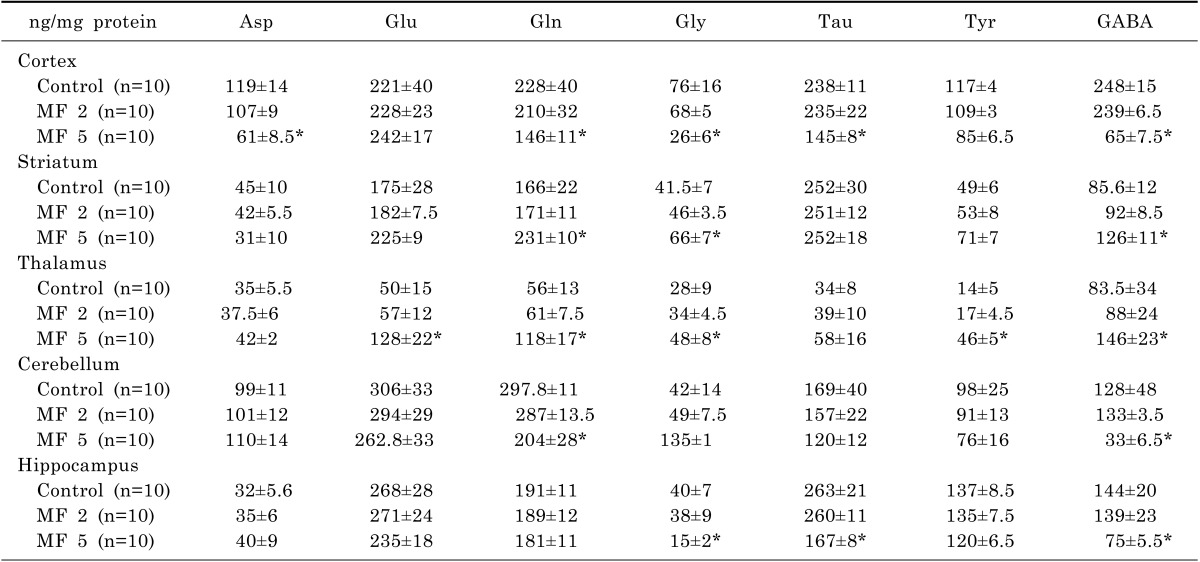
In contrast, ELF-MF induced the reduction of aspartic acid (Asp), Gln, Gly, Tyr and GABA in the cortex (p<0.05). Gln and GABA were significantly decreased by ELF-MF in the cerebellum (p<0.05). In the hippocampus, Gly, Tyr and GABA were also decreased by ELF-MF (p<0.05).
Determination of nitric oxide
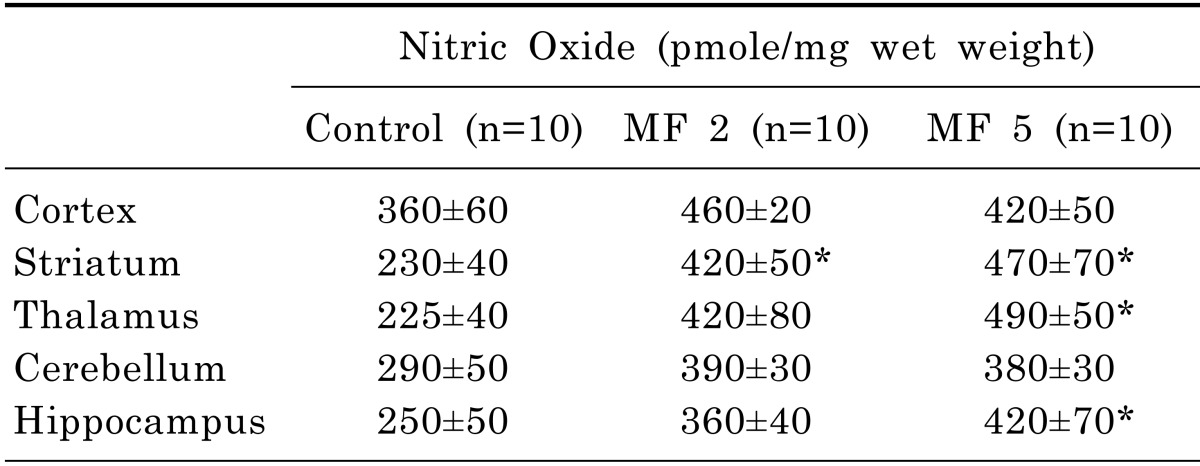
The exposure to 2.0 mT ELF-MF for 5 days alters nitric oxide (NO) concentrations significantly in the striatum, thalamus and hippocampus (p<0.05; Table 3). In the striatum, significant difference (p<0.05) was also observed in 2 day exposure of ELF-MF. In the cortex and cerebellum, no changes were obtained the groups exposed to 2.0 mT ELF-MF for 2 or 5 days.
Go to :
DISCUSSION
The main outcome in this study is that ELF-MF induces significant changes in the level of various neurotransmitters in the brain. The exposure of ELF-MF for 2 or 5 days altered significantly the level of NE and its metabolite, VMA in the striatum, thalamus, cerebellum and hippocampus. There were significant increases in 5-HT and HIAA in the striatum and thalamus or hippocampus exposed to ELF-MF for 5 days. The concentration of DA was significantly increased in the thalamus after the exposure to ELF-MF for 5 days. ELF-MF increased the levels of amino acid neurotransmitters such as Gln, Gly and GABA in the striatum and thalamus, whereas it decreased the levels in the cortex, cerebellum and hippocampus. NO concentration was significantly increased in the striatum, thalamus and hippocampus by ELF-MF.
One of the frequently observed biological effects of ELF-MF is the inhibition of production and secretion of a pineal hormone, melatonin, in pineal gland [25,26]. The enzyme involved in the synthesis of melatonin, serotonin-N-acetyltransferase (NAT), is blocked by ELF-MF [26]. It is well known that the concentration of melatonin in pineal gland is mediated via the adrenergic regulation, in which a kind of electromagnetic fields, light, depresses the release of NE in pinealocyte in daytime [27,28]. This means that ELF-MF may be related to regulation of neurotransmitters in animal brains. The modulation of receptor binding by ELF-MF was also confirmed by many reporters, suggesting that ELF-MF is involved in numerous CNS physiological function (learning and memory, sleep, temperature regulation, sexual behavior, etc.) and various pathophysiological disorders (anxiety, mood disorders, depression aggressiveness) [29,30,31]. Other workers suggested that MF desensitized membrane receptor in brain [32]. They discussed that this effect may explain some of the clinical influence observed or described after exposure to ELF-MF. From these works, it is speculated that ELF-MF exposure may act on CNS systems [4].
The neurotransmitters we assayed are the major neuronal neurotransmitters in the mammalian CNS, playing roles in a variety of physiological and pathological processes. Our results accumulate the evidences that ELF-MF seems to influence the physiology and pathophysiology in brain. The change in the level of neurotransmitters by ELF-MF probably acts on the neuronal communication in the brain because the major affected neurotransmitters are involved in psychological, psychiatric and motor symptoms. For example, biogenic amine neurotransmitters are related to sleep, alertness, stress response, addictive behaviors, anxiety and mood [33,34]. The observation that the elevation of 5-HT and its metabolite, HIAA, were all produced by ELF-MF means that ELF-MF might lead to physiological changes, particularly in the mood disorders.
Amino acid neurotransmitters are associated with excitatory and inhibitory neurotransmission [35]. In our amino acid study, ELF-MF exposure to rat increased or decreased the brain tissue concentration of amino acid neurotransmitters. The changes of tissue concentration by ELF-MF were not in agreement between brain tissue areas. For example, inhibitory amino acid neurotransmitter, GABA that was elevated in thalamus and striatum, was significantly decreased in cortex, cerebellum and hippocampus. This respectively different response in each brain tissue also appeared in excitatory amino acid neurotransmitter, glutamate or glutamine. Although it is difficult to point out one mechanism of how ELF-MF acts on CNS, the importance is that ELF-MF exposure can modulates the physiological neurotransmission in inhibitory or excitatory amino acids.
ELF-MF also induced the increment of NO levels in several brain regions. Since the first demonstration that endogenous NO modulates the release of acetylcholine in the basal forebrain, the effect of this gaseous messenger on the release of neurotransmitters has been extensively investigated. It has been shown that the release of various neurotransmitters, such as dopamine, inhibitory and excitatory amino acids, may either be increased or decreased by NO [36,37,38]. In the medial preoptic area and the striatum, serotonin release is enhanced by the NO precursor, L-arginine, and NO donors, respectively [38]. Although we do not speculate the interaction between ELF-MF and neuronal cells, it is inferred that ELF-MF may act on them through the unexplainable stimulation to neuronal cells resulting the alteration of the neurotransmitter levels.
From our results, we can speculate that there is possible relationship between ELF-MF and neurotransmitters. The present results may present a hint in favor of the existence of a cause-effect relationship between ELF-MF exposure and neurological disorders. Of course, a pre-requisite for being extrapolation of the present results to humans would be normalization for body mass of the ELF-MF effects. Moreover, it must be pointed out that at present, the mechanisms by which ELF-MF might interfere with the functions of neurotransmitters and receptors of the CNS are unknown.
Go to :
 XML Download
XML Download