

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
The increased use of electric power for domestic and industrial appliances in homes and in the workplace has resulted in increased exposure to 60-Hz extremely low frequency magnetic fields (ELF-MFs). Many studies have reported possible associations between ELF-MF exposure and an increased risk of cancer, genomic instability, and reproductive dysfunction [1,2,3,4]. However, the association between ELF-MFs and cancer remains controversial since different results have been obtained depending on the type of cancer studied; moreover, methodological problems have been identified in some of the studies [5,6]. Furthermore, one in vitro study found that a 50/60 Hz ELF-MF was unable to induce mutations [7]. Therefore, it has been hypothesized that ELF-MFs might not directly induce DNA damage; rather, they may affect cellular responses in cells already damaged by ionizing radiation (IR) or reactive oxygen species (ROS). Indeed, due to the suspected link between ELF-MFs and leukemia, a number of studies have investigated whether cell cycle distribution in mammalian cells is affected by exposure to ELF-MFs. These studies have focused on the effects of ELF-MFs on cell cycle-related end-points [8,9,10,11]. These studies concluded that not all cell types respond to MFs, yet it has been difficult to draw any additional conclusions regarding the necessary field characteristics to affect cells.
Importantly, few studies have focused on the cellular proliferation response to ELF-MFs; moreover, most of these studies used magnetic flux densities greater than 100 µT. These values were previously proposed by the International Commission on Non-Ionizing Radiation Protection (ICNIRP) [12] for the protection of workers and the general public, respectively, against exposure to 50-Hz MFs. However, ICNIRP guideline for workers was recently revised to 1 mT [13]. These studies have yielded conflicting results, including proliferative effects [14,15], antiproliferative responses [16,17] and no effect [18]. These inconsistencies have been proposed to be due to the nature of ELF-MF-induced biological responses, which are themselves influenced by a number of biological factors such as genetic characteristics and cellular differentiation status, in addition to physical factors such as field parameters and the chronological pattern of exposure [4,19].
Previous reports have suggested that ELF-MFs can act as promoters or cocarcinogenic factors in initiated cells. The World Health Organization (WHO) has also suggested that the primary research priority with regard to ELF-EMFs is to determine the co-carcinogenic effects of ELF-MFs in an in vitro experimental system [20].
We previously demonstrated that ELF-MFs given in combination with IR, hydrogen peroxide (H2O2), or overexpressed c-Myc (cellular myelocytomatosis oncogene) exerted no synergistic effects on transformation activity in mouse embryonic fibroblast NIH3T3 cells [21]. In another study, we also determined whether ELF-MFs affected the formation of micronuclei (MN) when given in combination with IR, H2O2, or c-Myc overexpression in NIH3T3 cells or WI38 human lung fibroblasts [22]. These studies did not reveal any effects on either transformation activity, or MN formation by 1 mT ELF-MFs, when given in combination with IR, H2O2, or c-Myc overexpression, in multiple cell lines. However, when a more sensitive method (detection of γ-H2AX) was used to examine the extent of DNA damage inflicted by ELF-MFs in combination with IR or H2O2 in non-tumorigenic cell systems, such as WI38 or L132 human lung epithelial cells, ELF-MFs increased the levels of γ-H2AX expression and the numbers of γ-H2AX foci. Of particular note, the intensity of the ELF-MFs was increased to 2 mT in this study, and cells were exposed to 60-Hz, 2 mT ELF-MFs. Moreover, exposure to a 2 mT ELF-MF potentiated the expression of γ-H2AX and the number of γ-H2AX foci when the field was given in combination with IR. However, the ELF-MF did not affect these responses when it was given in combination with H2O2 [23]. In this study, we examined the cell cycle distributions of nontumorigenic L132 cells exposed to ELF-MFs, with a particular focus on G2/M arrest; we also examined the extent of aneuploidy, a common marker of genomic instability that may play an important role in the early stages of cancer progression [24].
METHODS
ELF-MF exposure
The apparatus used to generate the ELF-MFs was designed and constructed by the Korea Electrotechnology Research Institute (KERI, Changwon, Korea) [21].
Cell culture
L132 human lung epithelial cells were purchased from the American Type Culture Collection (ATCC, Manassas, VA, USA) and maintained in Dulbecco's Modified Eagle's Medium (DMEM, GIBCO-Invitrogen, Paisley, Scotland, UK) or Roswell Park Memorial Institute medium (RPMI, GIBCO-Invitrogen). Medium was supplemented with 10% fetal bovine serum (Hyclone-Thermo Fisher Scientific, Logan, UT, USA). Cells were maintained at 37℃ in a humidified incubator in an atmosphere of 95% air and 5% CO2.
Exposure to ELF-MFs, IR, and H2O2
Cells were seeded in 60-mm cell culture dishes, placed in the exposure chamber, and subsequently exposed to a 60-Hz ELF-MF for 9 h. The temperature in the chamber was maintained at 37±0.3℃ using a circulating water system, and the temperature of the culture medium was monitored at 2 h intervals throughout the duration of the exposure. As positive controls, cells were exposed to various gamma radiation doses (0~2 Gy in a single dose) generated by a 137Cs gamma-ray source (MDS Nordion, Ottawa, ON, Canada) at a dose rate of 5 Gy/min. For H2O2 (Sigma, St. Louis, MO, USA) exposure, cells in 60-mm cell culture dishes were treated with H2O2 (0~0.05 mM) for 9 h. The following treatments were used in the combination experiments: exposure to an ELF-MF (1 mT or 2 mT) for 9 h, immediately after irradiation with gamma rays; exposure to an ELF-MF for 9 h in the presence of H2O2; and sham exposure in the cell culture incubator.
Flow cytometry analysis
Cell cycle distributions were analyzed using propidium iodide (Sigma, St. Louis, MO, USA) staining followed by flow cytometry. Cells were seeded at a density of 2.5×105 cells/dish in 60-mm cell culture dishes and incubated overnight. Cells were then collected by trypsinization and harvested by centrifugation at 1300 rpm for 3 min. Next, cells were fixed with 70% cold ethanol at -20℃ overnight, washed with phosphate-buffered saline (PBS), treated with RNaseA (1 mg/ml; Sigma, St. Louis, Missouri, USA), and stained with propidium iodide (50 µg/ml). Samples were analyzed using a FACSCalibur flow cytometer (BD Biosciences, San Jose, CA, USA); data were analyzed with BD CellQuest Pro Software (BD Biosciences).
RESULTS
Induction of aneuploidy by IR and H2O2
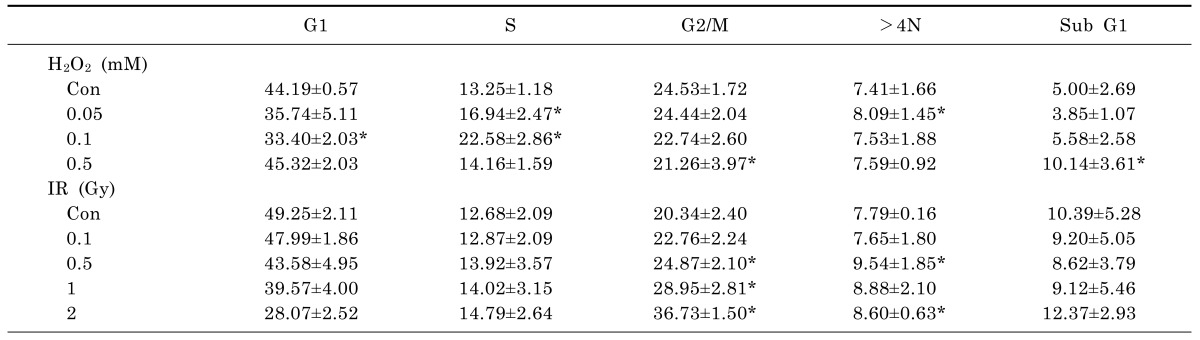
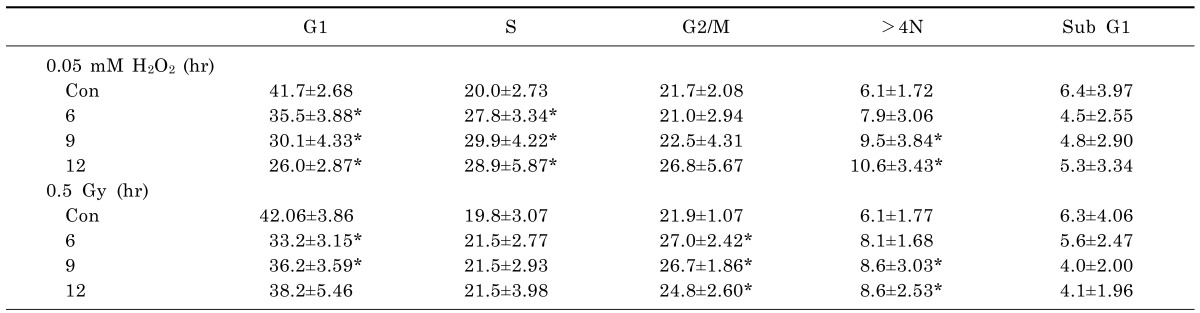
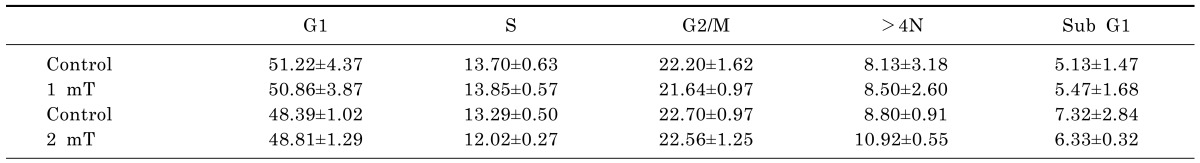
To determine the appropriate H2O2 concentrations and IR doses to use in combination with ELF-MFs, non-tumorigenic L132 human lung epithelial cells were treated with various concentrations of H2O2 or various doses of IR. After 9 h of culture, the distribution of cells in various cell cycle stages, including subG1 (induction of apoptosis), G2/M (before aneuploidy formation), and >4 N (aneuploidy production), as well as G1 and S was examined by flow cytometry (Table 1). Compared with control cells, cells treated with H2O2 or IR alone exhibited significantly altered levels of aneuploidy. The strongest effects on aneuploidy were observed after treatment with 0.05 mM H2O2 or exposure to 0.5 Gy IR; importantly, none of these treatments altered the proportion of cells in subG1, indicating the absence of apoptotic cell death. Kinetic analysis of the induction of aneuploidy by 0.05 mM H2O2 or 0.5 Gy IR revealed that aneuploid cells were observed after 9 h of either treatment (Table 2). The effects of exposure to ELF-MFs (1 mT or 2 mT for 9 h) on aneuploidy induction in L132 cells were also examined. At strengths of 1 mT or 2 mT, ELF-MFs did not alter the distributions of G2/M-arrested cells or aneuploid cells. Similarly, ELF-MFs did not induce apoptosis (Table 3). In subsequent experiments, either 0.05 mM H2O2 or 0.5 Gy IR was used, since these doses corresponded to the minimum doses of IR or H2O2, respectively, required to affect cell aneuploidy. At least four biological replicates were performed for each experiment; representative results are shown.
ELF-MFs do not affect H2O2-induced or IR-induced aneuploidy
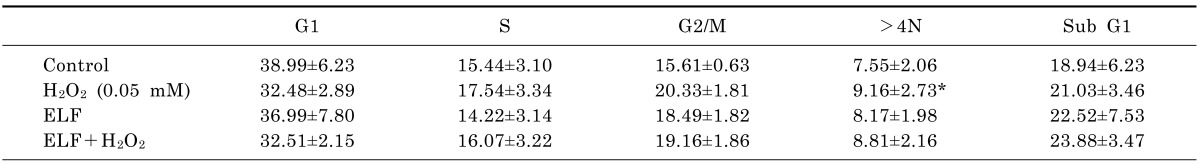
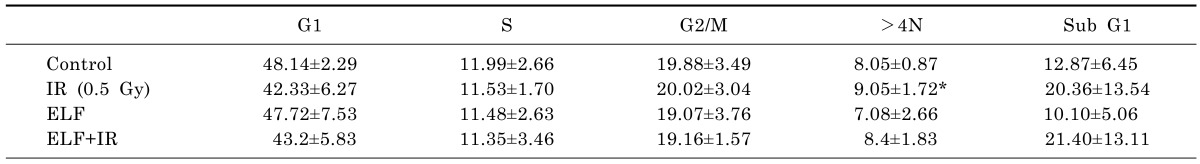
To determine whether ELF-MFs in combination with H2O2 or IR could affect aneuploidy formation, L132 cells were treated with either 0.05 mM H2O2 or 0.5 Gy IR and simultaneously exposed to an ELF-MF (2 mT) for 9 h. Subsequently, the percentages of G2/M-arrested cells and aneuploid cells were examined by flow cytometry. Interestingly, neither H2O2-induced nor IR-induced changes in the proportions of >4N cells were affected by treatment with a 2 mT ELF-MF. Moreover, neither H2O2 nor IR treatment affected the population of subG1 cells compared with control cells; this result was not affected by simultaneous treatment with an ELF-MF. These results indicate that ELF-MFs do not affect H2O2-induced or IR-induced aneuploidy, at least in non-tumorigenic cell lines such as L132 cells. At least five biological replicates were performed for each experiment, with each yielding similar results (Tables 4 and 5). In addition, when we performed multinucleated cells using microscopical visualization method, similar results were obtained (data not shown).
DISCUSSION
The results presented here show that 9 h of exposure to a 2 mT ELF-MF does not affect either IR-induced or H2O2-induced G2/M arrest (which occurs before aneuploidy); the same treatment also does not affect the extent of aneuploidy, one of the initial alterations that leads to genomic instability. We chose non-tumorigenic L132 human lung epithelial cells for this study because cancer cells usually show a high incidence of aneuploidy. In this study, the magnetic flux intensities (1 mT or 2 mT) were selected based on Korean exposure guidelines; moreover, the selected magnetic flux intensities were equal to or up to 10-fold higher than the reference levels proposed by the ICNIRP [13]. These reference levels were 1 mT for occupational exposure and 200 µM for exposure of the general public.
Aneuploidy is a hallmark of tumor cells [25]; more than 90% of all solid tumor cells have been shown to be aneuploid [26]. Aneuploidy has been reported to precede and to segregate with chemical carcinogenesis [27]. Thus, aneuploidy is believed to play a causal role in tumorigenesis [28]. The precise contribution of aneuploidy to tumorigenesis is not yet fully understood; however, insights into the mechanisms of aneuploidy will surely be valuable for better understanding the mechanisms of tumorigenesis. Aneuploidy is a condition in which the chromosome number is not an exact multiple of the number that is characteristic of a particular species. Having an extra or a missing chromosome is one common cause of genetic disorders. Some cancer cells have been shown to have abnormal numbers of chromosomes [29]. Moreover, some chemicals that prevent the formation of the spindle apparatus during mitosis generate whole chromatids, which are excluded from nuclei. This phenomenon leads to the formation of multinucleated, aneuploid cells, in which each nucleus contains a different number of chromosomes [30]. Genomic instability is defined as an increased acquisition rate of alterations in the genome [31]. The most notable types of genomic instability in cells involve aneuploidy [32], gene deletion [33], and chromosomal abnormalities [34].
Since Wertheimer and Leeper (1979) reported a correlation between ELF-MF exposure and childhood cancer, many epidemiological studies have investigated whether ELF-MFs are associated with an increased risk of cancer. It is generally believed that ELF-MFs alone do not produce sufficient energy to induce DNA or chromosomal damage [35], and that exposure to known carcinogens is probably part of the link between cancer and ELF-MFs [36,37]. In accordance with this hypothesis, we found that exposure to either a 1 mT or a 2 mT ELF-MF alone did not induce aneuploidy in human lung epithelial cells.
Treatment with either IR or H2O2 is well established to promote both cellular aneuploidy [38,39] and genomic instability [31,40]. We selected IR as a representative DNA-damaging agent commonly found in the environment, whereas H2O2 was employed as a ROS factor. In the present study, IR and H2O2 were positive controls used to demonstrate the responsiveness of cells to known tumor-initiating agents. We found that a 0.5 Gy dose of IR or treatment with 0.05 mM H2O2 for 9 h resulted in the highest proportion of aneuploid cells for each control. Higher doses of radiation and higher concentrations of H2O2 did not increase the proportions of aneuploid cells; this result may have been due to cell death, although the populations of subG1 cells were not significantly increased in these experiments. We also examined the effects of these controls on G2/M phase arrest, which usually occurs in cells with defective mitosis and cytokinesis before aneuploidy is observed [41]. We found that IR increased the extent of G2/M phase arrest and the induction of aneuploidy (with the strongest effect observed on G2/M arrest); in contrast, H2O2 only induced aneuploidy, suggesting that IR and H2O2 contribute to aneuploidy via different mechanisms. While IR-induced genomic instability is generally attributed to DNA damage immediately after exposure, mistakes in DNA damage repair, or infidelity in DNA replication [42], several studies have suggested that the genetic consequences of irradiation might be delayed and only appear in the descendants of irradiated cells after several generations [33,43,44,45]. These delayed effects are later manifested as reproductive death, chromosomal instability, and mutagenesis [34,46]. Thus, IR-induced genomic instability may lead to a variety of cellular changes including chromosomal instability, which is a serious form of genomic instability; moreover, IR-induced genomic instability may also play an important role in the early stages of cancer progression [24]. In contrast, ROS such as H2O2 are themselves mutagenic and are capable of producing DNA strand breaks, chromosomal deletions, chromosomal rearrangements, and altered expression of proto-oncogenes and tumor suppressor genes [47]. Furthermore, ROS may interact with and modify cellular proteins and lipids, resulting in alterations in cellular functions such as proliferation, development, senescence, and apoptosis [48]. Therefore, IR and ROS may induce aneuploidy via different mechanisms. In a previous study using bleomycin as an initiator followed by exposure to a 0.8 mT ELF-MF, the frequencies of MN and aneuploidy were significantly increased [49]. However, we found that the combination of an ELF-MF with either IR or H2O2 did not induce any more aneuploidy than was induced by each stimulus alone.
Our previous studies showed that exposure to both a 1 mT ELF-MF and IR for a maximum of 24 h did not enhance MN formation or comet tail production in several cell lines [22,50]. However, exposure to a 2 mT ELF-MF (but not a 1 mT ELF-MF) in combination with IR significantly increased the level of γ-H2AX. In contrast, exposure to a 2 mT ELF-MF in combination with H2O2 did not significantly alter the level of γ-H2AX [23]. We did not observe any alterations in G2/M phase arrest, aneuploidy patterns, or cell death upon exposure to a 2 mT ELF-MF in the present study. Therefore, further studies of the effects on MN formation and comet tail production by exposure to a 2 mT ELF-MF in combination with IR will be important for confirming the genotoxic effects of ELF-MFs in combination with IR.
 XML Download
XML Download