

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
P19 mouse embryonal carcinoma cells (P19 cells) can differentiate into neuron-like cells when induced in the presence of retinoic acid [1]. Since neuronally differentiated P19 cells express many kinds of neuron-related molecules including functionally active N-methyl-D-aspartate (NMDA) receptors [2,3], which play roles in the processes of neuronal development and in the formation of synaptic plasticity, P19 cells have been used as a model system to study the function of NMDA receptors present in the central nervous system (CNS).
Nogo receptor 1 (NgR1), the receptor for neurite outgrowth inhibitors, such as Nogo-A, myelin-associated glycoprotein (MAG) and oligodendrocyte myelin glycoprotein (OMgp), mediates axonal growth inhibition in the CNS, and was more recently reported to participate in other functions including neuronal differentiation and synapse formation [4]. NgR1 and Nogo A are localized in the synapses of neurons and appear to participate in the regulation of long-term potentiation (LTP) in adult CNS tissue [5,6,7].
To study the function of nogo receptors and their ligands, cell culture systems using human cancer cells, murine embryonic stem cells or rat neural progenitor cells have been exploited. Some signaling pathways stimulated by activation of the nogo receptors expressed in those systems have been elucidated [8,9,10]. However, these cell culture systems do not fully represent the complex neural system of the CNS. Thus, it is necessary to find and develop diverse cell culture models that express Nogo receptors and related signaling molecules to further the understanding of the Nogo system. In the present study, we report for the first time that NgR1 is expressed in neuron-like P19 cells differentiated by all trans-retinoic acid and can transmit a signal when stimulated by its ligand.
METHODS
Cells and reagents
P19 mouse embryonal carcinoma cells and SH-SY5Y human neuroblastoma cells (SH-SY5Y cells) were obtained from American Type Culture Collection (Manassas, VA). Antibody against NgR1 and Nogo-P4 and NEP1-40 peptides were purchased from Alpha Diagnostic International (San Antonio, TX). Antibodies against STAT3 (signal transducer and activator of transcription 3), phosphor-STAT3, mTOR (mammalian target of rapamycin), and phosphor-mTOR were from Cell Signaling Technology (Danver, MA). Neurobasal medium, α-MEM, and N2 supplement were obtained from Life Technologies (Carlsbad, CA). All other reagents were from Sigma-Aldrich (St. Louis, MO).
Induction of differentiation of P19 and SH-SY5Y cells
P19 cells were maintained in α-MEM supplemented with 10% heat-inactivated fetal bovine serum (FBS). Differentiation of P19 cells into neuronal P19 cells were induced as described previously [11]. Briefly, to induce neuronal differentiation of P19 cells, the cells were stimulated to form embryonic bodies in bacteriological-grade Petri dishes by addition of 0.5 µM all-trans retinoic acid at a density of 1×106 cells in α-MEM supplemented with 5% heat-inactivated FBS. After 4 days of incubation, the cellular aggregates were collected, trypsinized, and transferred into poly-d-lysine coated cell culture dishes. The cells were cultured in neurobasal medium containing N2 supplement for an additional 5 days. Before treating the cells with Nogo-P4 and/or NEP1-40 peptides for 30 min, the differentiated cells were incubated for 16 h in the absence of N2 supplement. SH-SY5Y cells were maintained in Ham's F12 medium with 10% FBS. Neuronal differentiation of SH-SY5Y cells were induced by stimulating the cells with 10 µM all trans-retinoic acid alone for 5 days or by exposing the cells sequentially to 10 µM all trans-retinoic acid for 5 days and 25 ng/ml brain-derived neurotrophic factor (BDNF) for an additional 24 h [8].
RNA extraction and Reverse Transcription-PCR
RNA was extracted from cells with RNeasy kit (Qiagen, Valencia, CA) according to the manufacturer's protocol. Approximately 1 µg of total RNA was used to prepare cDNA using Qiagen one step RT-PCR kit (Qiagen). The nucleotide sequences of the primers for NgR1, NMDA receptors and β-actin are listed in Table 1. The cycles of PCR reactions were determined on 25~35 cycles to allow semiquantitative comparisons of cDNAs. The RT-PCR products were electrophoresed on a 1 to 3% agarose gel and visualized via staining with ethidium bromide.
Western immunoblot analysis
Total protein extracts were prepared by lysing cells in lysis buffer (50 mM Tris-Cl [pH 7.4], 150 mM NaCl, 1 mM EGTA, 5 mM EDTA, 0.5% Triton X-100, 0.25% sodium deoxycholate, 1 mM sodium fluoride, 1 mM sodium orthovanadate, 5 µg/ml leupeptin, 0.2 mM phenylmethylsulfonyl fluoride, 10 µg/ml aprotinin, and 0.5 mM dithiothreitol). Protein extracts were resolved by 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred to nitrocellulose membranes. The membranes were incubated with blocking buffer (Tris-buffered saline containing 0.2% Tween-20 and 5% nonfat dried milk) and probed with the indicated primary antibody. After washing, membranes were probed with horseradish peroxidase-conjugated secondary antibody. Detection was performed using an enhanced chemiluminescent protein detection system (Amersham Biosciences, Little Chalfont, United Kingdom) and exposure was taken using X-ray film. The band density was quantified by ImageJ software.
RESULTS
Expression of NgR1 in P19 cells differentiated by all trans-retinoic acid
Parental P19 cells did not produce any branches that could be seen as neurites when grown in normal maintenance medium (Fig. 1A). P19 cells exposed to 0.5 µM all trans-retinoic acid in bacteriological grade Petri dishes formed embryonic bodies suspended in the medium. After single cells prepared from the embryonic bodies were transferred to poly-d-lysine coated culture dishes containing N2-supplemented culture medium, the cells started producing branches and then formed extensive amounts of neurites (Fig. 1B). Since neuronally differentiated P19 cells have characteristics of CNS neurons [3] and NgR1 is also expressed mainly in the CNS [12], the possibility that P19 cells differentiated by all trans-retinoic acid could express NgR1 was examined. The expression of mRNA and protein of NgR1 was measured by semiquantitative RT-PCR and western immunoblotting analysis, respectively. Interestingly, as shown in Fig. 2, the mRNA and protein of NgR1 was expressed in differentiated P19 cells, but was not detected in the parental P19 cells and the embryonic bodies formed from P19 cells in the presence of all trans-retinoic acid.
Expression of NMDA receptors in neuronally differentiated P19 and SH-SY5Y cells
NMDA receptors are mainly expressed in CNS neurons. To determine whether mRNA of the NR1, NR2A, NR2B, NR2C, and NR2D subunits of the NMDA receptor are expressed in neuronally differentiated P19 cells and SH-SY5Y cells, semiquantitative RT-PCR analysis was performed (Fig. 3). The parental P19 cells and embryonic bodies expressed mRNA of NR1 and NR2A at a detectable level, but the expression of mRNA of NR2B, NR2C, and NR2D was hardly detected. Neuronally differentiated P19 cells abundantly expressed all of the NMDA receptor subtypes up to the fifth day of differentiation (Fig. 3A). SH-SY5Y cells, which express the NgR1 mRNA and protein when differentiated neuronally [8], expressed only some mRNAs of the human NMDA receptor subtypes (Fig. 3B). In all experimental groups of SH-SY5 cells, the mRNA of NR1, NR2C, and NR2D were expressed at a high level, but those of NR2A and 2B were barely detectable.
Activation of STAT3 by NgR1 expressed by neuronally differentiated P19 cells
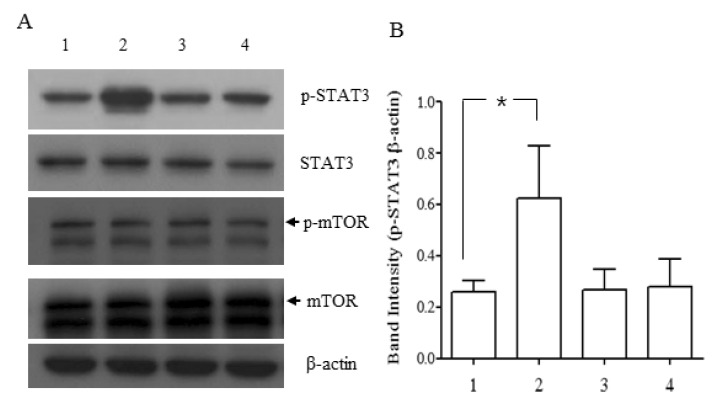
Two recent reports showed that NgR1 expressed in murine embryoid bodies or in astroglial lineage cells differentiated from rat stem/progenitor cells (NPCs) transmitted its signal via STAT3 when activated by their ligand [9,10]. To determine whether NgR1 expressed in neuronally differentiated P19 cells would be functional, P19 cells differentiated for 5 days were exposed to Nogo-P4 peptide, an active segment of Nogo-66. As shown in Fig. 4, Nogo-P4 peptide increased the phosphorylation of STAT3 and co-treatment of NEP1-40, a specific Nogo-66 receptor antagonist, with Nogo-P4 returned the phosphorylation status of STAT3 to its basal level. Phosphorylation of mTOR was increased in rat NPCs by treatment with a Nogo receptor agonist. But, in the present study, the level of phosphorylation of mTOR was not affected in the presence and absence of Nogo-P4 peptide.
DISCUSSION
The presence of NgR1 was discovered in the process of investigating the reason why CNS neurons have very limited ability to repair and regenerate axons after injury. Many studies have suggested that CNS myelin sheath contains components responsible for inhibiting neurite outgrowth of CNS neurons. The nogo gene, which expresses Nogo-A, a major neurite outgrowth inhibitor, was identified at the same time by three research groups [13,14,15]. NgR1 that had a high affinity with Nogo-66, an active segment of Nogo-A, was discovered soon after the identification of the nogo gene [16]. Further studies showed that NgR1 also binds and interacts with OMgp and MAG [17,18], although MAG binds with higher affinity to Nogo receptor 2 (NgR2) [19]. Although P19 cells are mouse cells, they have been actively exploited to investigate characteristics of CNS neurons because a limited number of cell culture models can serve as surrogates for CNS neurons of a human and, in many cases, they do not possess characteristics that investigators need. Neuronally differentiated P19 cells express various kinds of receptors and ion channels present in normal neurons, such as NMDA receptors, AKPA/kainate receptors, nicotinic and mucarinic receptors, and calcium channels [20]. However, the presence of NgR1 in P19 cells has not been examined so far, although it is a component that is abundantly expressed in CNS neurons.
The present study shows for the first time that mRNA and protein of NgR1 are expressed in neuronally differentiated P19 cells. The mRNA of NgR1 was weakly expressed in parental P19 cells before differentiation and the embryonic bodies, but the expression of NgR1 protein was not detected. These results suggest that the NgR1 protein may play some roles after initiation of differentiation.
The presence and function of NMDA receptors in neuronally differentiated P19 cells have been proven and studied. In addition to the involvement of NMDA receptors in neurotoxicity, they have been reported to play a crucial role in controlling synaptic plasticity and memory formation [21,22]. In the present study, the expression of mRNAs of NMDA receptor subtypes NR1 and NR2A-NR2D was clearly observed during the period of differentiation, but the expression of a part of mRNAs of NMDA receptor subtypes was not detected in parental P19 cells and embryonic bodies. These present and previous results bolster the view that neuronally-differentiated P19 cells express functionally active NMDA receptors. Recent studies also suggest that NgR1 and Nogo A might play roles in synaptic plasticity [4]. The fact that NMDA receptors and NgR1 could play in synaptic plasticity raises the need to investigate the possibility of an interaction between NMDA receptors and NgR1. A recent report showed that Nogo A regulated the level of expression of AMPA and NMDA glutamate receptor subunits in hippocampal neurons [23]. The neuronally differentiated P19 cells might provide an efficient in vitro model system to study any relation between signaling events of NMDA receptors and NgR1 because these cells express both types of receptors.
Recently, there was a report that SH-SY5Y human neuroblastoma cells could express functionally active NgR1 when differentiated by a combined treatment of brain-derived neurotrophic factor (BDNF) and retinoic acid [8]. In our study, regardless of the status of differentiation, SH-SY5Y cells clearly expressed mRNA of NR1, NR2C, and NR2D with very weak expression of NR2A mRNA. A previous study showed that SH-SY5Y cells only expressed NR1 mRNA, regardless of differentiation status [24]. This discrepancy might be due to the differences of the PCR primers. Although it is controversial, it appears that SH-SY5Y cells do not express functional NMDA receptors in view of the studies conducted by others until now. Thus, in terms of studying the relation between signaling events of NgR1 and NMDA receptors, neuronally differentiated P19 cells seem to be equipped with better tools. A different cell culture system using embryonic stem cells that express functional NgR1 was reported [9], but the system has some limitations, such as difficulty in obtaining and maintaining embryonic stem cells.
The RhoA/RHO kinase pathway mediates neurite outgrowth and axonal regeneration exerted by the activation of NgR1 by Nogo-66 [25,26]. In addition to this well-known RhoA/RHO kinase pathway, two other pathways including STAT3 and mTOR were suggested to be activated in rat neural progenitor cells when treated with Nogo-66 [10]. However, in our model, phosphorylation of STAT3 was increased by the treatment of Nogo-P4 peptide, but that of mTOR was not affected. These results show that NgR1 expressed in the neuronally differentiated P19 cells was functionally working in terms of signal transduction.
Since NgR1 alone cannot produce signaling events owing to absence of a transmembrane domain and a cytoplasmic domain, it needs binding partners, such as LINGO1, p75, and TROY [4]. It is believed that neuronally differentiated P19 cells provide a good in vitro experimental model to study the presence and role of these Nogo binding partners and signaling molecules activated by Nogo-66. Recently, Nogo and NgR1 are also being studied for their possible involvement in CNS disorders or diseases, such as Alzheimer's disease, schizophrenia and multiple sclerosis [27,28,29]. We also believe that this P19 cell model could be an efficient tool to elucidate whether signaling events of Nogo and NgR1 interact with each other to perform any physiological functions, such as LTP or LTD (long-term depression) or participate in pathologic conditions of the CNS mentioned above.




 XML Download
XML Download