

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Hydroxycinnamic acids have been reported to possess a wide variety of biological properties such as antioxidant, anti-inflammatory, anti-tumor, and neuroprotective activities [1-4]. We demonstrated that 3,4,5-trihydroxycinnamic acid, a derivative of hydroxycinnamic acid, significantly attenuates LPS-induced inflammatory responses through the suppression of NF-κB transcription and the activation of Nrf2 signaling pathway in BV2 microglial cells [5,6]. In addition, ester derivatives of hydroxycinnamic acids have been also reported to exhibit numerous biological properties [7]. Caffeic acid phenylethyl ester (CAPE) inhibits cytokine-mediated NF-κB activation through the activation of Nrf2 signaling pathway in macrophage cells [8]. Recently, we also observed anti-inflammatory activity of methyl p-hydroxycinnamate, a methyl ester of hydroxycinnamic acid, in LPS-induced RAW264.7 macroglial cells. Methyl p-hydroxycinnamate significantly attenuated LPS-induced inflammatory responses (Under review). Given the facts that methyl p-hydroxycinnamate, a methyl ester of hydroxycinnamic acid, possesses a variety of biological activities such as anti-tumor [9], anti-oxidant [10], depigmenting [11], and anti-adipogenic [12] activities, 1-p-coumaroyl β-D-glucose (CG), a glucose ester of hydroxycinnamic acid, has a potential to exhibit biological activities. However, the anti-inflammatory property of CG has not been clearly examined. CG has been reported to be present in berry fruits including strawberries (Fragaria x ananassa) [13]. CG, used in the present study, was isolated from the bark of Salix hulteni [14].
Macrophages play an major role in inflammation and mobilization of the host defense against bacterial infection [15]. However aberrant activation of macrophages has been also reported to play a pathogenic role in various inflammatory disorders including sepsis [16]. In pathogenic conditions, abnormally activated macrophages produce excessive amount of a variety of pro-inflammatory mediators and cytokines through NF-κB-mediated pro-inflammatory transcription that eventually aggravates the inflammatory conditions [17]. Lipopolysaccharide (LPS), a component of the outer membrane of Gram-negative bacteria, is the most common cause of macrophage activation [18]. LPS-induced activation of macrophages has been reported to cause a wide range of responses including secretion of pro-inflammatory mediators, expression of adhesion molecules and coagulation factors, phagocytosis, and cytoskeletal rearrangement [19]. Therefore, suppression of aberrant activation of macrophage might be a valuable therapeutic target for the treatment of inflammatory disorders.
The objective of the present study was to examine the anti-inflammatory properties of CG and its underlying mechanism in LPS-stimulated RAW264.7 macrophage cells in order to provide a valuable pharmacological agent that could attenuate aberrant inflammatory responses in inflammation-related conditions.
Go to : 
METHODS
Reagents and cell culture
Bacterial lipopolysaccharide (LPS) from Escherichia coli serotype 055:B5 was purchased from Sigma-Aldrich (St. Louis, MO, USA). 1-p-Coumaryol β-D-glucoside (CG) was isolated and identified from the bark of Salix hulteni [14] (Fig. 1). The compound was dissolved in dimethyl sulfoxide (DMSO) and added to the cell culture at the desired concentrations. The macrophage RAW264.7 cells were maintained in Dulbecco's modified Eagle's medium (DMEM; Gibco BRL, Grand Island, NY, USA) containing 5% heat-inactivated fetal bovine serum and penicillin-streptomycin (Gibco BRL) at 37℃, 5% CO2. In all experiments, cells were incubated in the presence of the indicated concentration of CG before the addition of LPS (200 ng/ml).

Cell viability
Cell viability was determined by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. The RAW264.7 cells were seeded at 5×105 cells per well and incubated with CG at various concentrations for 24 hr at 37℃. After incubation, MTT (0.5 mg/ml in PBS) was added to each well, and the cells were incubated for 3 hr at 37℃ and 5% CO2. The resulting formazan crystals were dissolved in dimethyl sulfoxide (DMSO). Absorbance was determined at 540 nm. The results were expressed as a percentage of surviving cells over control cells.
Nitrite quantification assay
The production of NO was estimated by measuring the amount of nitrite, a stable metabolite of NO, using the Griess reagent as described [5]. After CG-pretreated RAW264.7 macrophage cells were stimulated with LPS in 12-well plates for 24 hr, 100 µl of the cell supernatant was mixed with an equal volume of Griess reagent. Light absorbance was read at 540 nm. The results were expressed as a percentage of released NO from LPS-stimulated RAW264.7 cells. To prepare a standard curve, sodium nitrite was used to prepare a standard curve.
Western blot analysis
The RAW264.7 macrophage cells were pretreated with CG for 1 hr and then stimulated with LPS. Cells were washed with PBS and lysed in PRO-PREP lysis buffer (iNtRON Biotechnology, Seongnam, Korea). Equal amounts of protein were separated on 10% SDS-polyacrylamide gel. Proteins were transferred to Hypond PVDF membrane (Amersham Biosciences, Piscataway, NJ, USA) and blocked in 5% skim milk in TBST for 1 hr at room temperature. Specific antibodies against iNOS, COX-2, extracellular signal-regulated kinase (ERK), phosphorylated (p)-ERK, p38, p-p38, c-Jun N-terminal kinase Akt, p-Akt (1 : 1,000; Cell Signaling Technology), IκB-α (1 : 1,000; Santa Cruz Biotechnology Inc), and β-actin (1 : 2,500; Sigma) were diluted in 5% skim milk. After thoroughly washing with TBST, horseradish peroxidase-conjugated secondary antibodies were applied. The blots were developed by the enhanced chemiluminescence detection (Amersham Biosciences).
ELISA assay for cytokines
The RAW264.7 macrophage cells were treated with CG in the absence or presence of LPS. After 24 hr incubation, TNF-α and IL-1β levels in culture media were quantified using monoclonal anti-TNF-α or IL-1β antibodies according to the manufacturer's instruction (R&D Systems).
Statistical analysis
All values shown in the figures are expressed as the mean±SD obtained from indicated numbers of independent experiments. Statistical significance was analyzed by two-tailed Student's t-test. Data with values of p<0.05 were considered as statistically significant. Single (* and #) and double (** and ##) marks represent statistical significance in p<0.05 and p<0.01, respectively.
Go to :
RESULTS
1-p-Coumaryol β-D-glucoside (CG) suppresses NO and PGE2 production in LPS-stimulated RAW264.7 macrophage cells
Excessive production of pro-inflammatory mediators such as NO and PGE2 is one of characteristic features of macrophage-mediated inflammatory conditions [5,20]. Therefore, inhibitory property of CG on NO and PGE2 production in LPS-stimulated RAW264.7 macrophage cells were examined. RWA264.7 cells were incubated with indicated concentrations of CG for 1 hr prior to LPS treatment and, for NO measurement, Griess reaction was used as an index for NO synthesis. CG significantly inhibited NO production in LPS-stimulated RAW264.7 cells (Fig. 2A) and PGE2 production in a concentration-dependent manner (Fig. 2B). To examine the anti-inflammatory magnitude of CG, which is a glucose ester of hydroxycinnamic acid, the inhibitory potency on LPS-induced NO production was compared with methyl p-hydroxycinnamate, a methyl ester of hydroxycinnamic acid. Inhibitory effect of CG on LPS-induced NO and PGE2 production was less potent than that of methyl p-hydroxycinnamate. In addition, CG showed no noticeable cytotoxicity in concentration ranges (Fig. 2C).
 | Fig. 2CG attenuated LPS-induced NO and PGE2 production in RAW264.7 macrophage cells. RAW264.7 cells were pretreated with various indicated concentrations of CG for 1 hr, then incubated with LPS (200 ng/ml) for 24 hrs. The concentrations of NO (A) and PGE2 (B) in the supernatants were measured by Griess and ELISA assay as described in the methods. CG exhibited a significant suppression of LPS-induced NO (A) and PGE2 (B) secretion in concentration-dependent manners. No significant cell death was observed at CG concentrations used in the present study (C). Methyl p-hydroxycinnamate (MH) was used as a reference compound compared with CG. The data are expressed as mean±SD for three (NO) or four (PGE2) independent experiments. *p<0.05 and **p<0.01 indicate statistically significant differences from treatments with LPS alone.
|
CG inhibits LPS-induced protein expression of iNOS and COX-2
As CG significantly suppressed LPS-induced NO and PGE2 production in RAW264.7 cells (Fig. 2), protein expression level of iNOS and COX-2 was examined. LPS resulted in noticeably increased expression of iNOS and COX-2 proteins and CG significantly attenuated protein expression levels in a concentration-dependent manner (Fig. 3), indicating that decreased production of NO and PGE2 is due to decreased expression of their responsible proteins, iNOS and COX-2, respectively, with CG treatment.
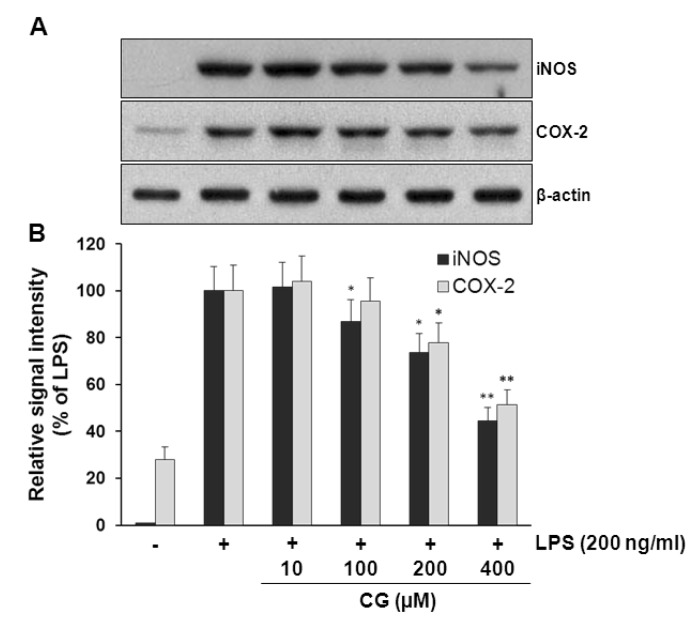 | Fig. 3CG inhibited LPS-induced iNOS and COX-2 protein expressions in RAW264.7 macrophage cells. (A) RAW264.7 cells were pretreated with indicated concentrations of CG for 1 hr, then incubated with LPS (200 ng/ml) for 24 hrs. The cell lysates were prepared and subjected to Western blotting analysis by using antibodies specific for iNOS and COX-2 as described in the methods. (B) The relative protein levels were quantified by scanning densitometry and normalized to β-actin. The data indicated that CG significantly decreased the LPS-induced overexpression of iNOS and COX-2 in a concentration-dependent manner. The images shown are representatives of three independent experiments that showed consistent results and the relative protein values are expressed as mean±SD for three experiments. *p<0.05 and **p<0.01 indicate statistically significant differences from treatments with LPS alone.
|
CG attenuates LPS-induced release of pro-inflammatory cytokines such as IL-1β and TNF-α
To examine the effects of CG on the extracellular secretion of pro-inflammatory cytokines such as IL-1β and TNF-α, secretion of these cytokines was measured using ELISA assay in LPS-stimulated RAW264.7 cells. LPS treatment resulted in excessive extracellular release of IL-1β and TNF-α in RWA 264.7 cells. CG significantly attenuated LPS-induced extracellular release of IL-1β and TNF-α in a concentration-dependent manner (Fig. 4).
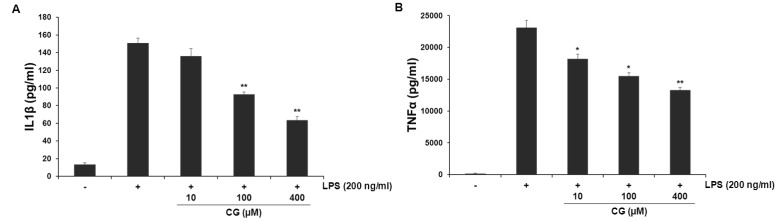 | Fig. 4CG suppressed the extracellular release of LPS-induced IL-1β and TNF-α cytokines in RAW264.7 macrophage cells. RAW264.7 cells were pretreated with indicated concentrations of CG for 1 hr, then incubated with LPS (200 ng/ml) for 24 hrs. The concentrations of IL-1β (A) and TNF-α (B) in collected cell culture media were measured by ELISA assay as described in the methods. CG meaningfully reduced LPS-stimulated both IL-1β and TNF-α cytokines in a concentration-dependent manner. The values are expressed as mean±SD for three independent experiments. *p<0.05 and **p<0.01 indicate statistically significant differences from treatments with LPS alone.
|
CG attenuates LPS-induced degradation of IκB
NF-κB has been widely demonstrated to be a major transcription factor of multiple pro-inflammatory genes. Cytosolic IκB negatively regulates NF-κB-mediated transcription by sequestering NF-κB in the cytoplasm given the fact that NF-κB in cytoplasm has to translocate to the nucleus for transcription of pro-inflammatory genes when NF-κB is freed due to proteasomal degradation of IκB [20, 21]. Once degradation of IκB In the present study, whether CG inhibits LPS-induced degradation of IκB was examined. LPS treatment resulted in the marked degradation of IκB and CG significantly attenuated LPS-induced degradation of IκB in a concentration-dependent manner (Fig. 5), suggesting that CG might subsequently suppress LPS-induced NF-κB transcription in RAW264.7 cells.
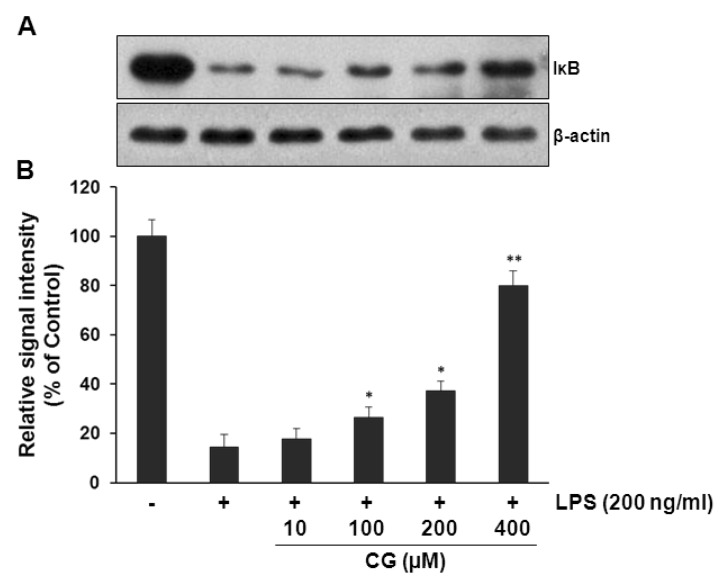 | Fig. 5CG significantly attenuated LPS-induced IκBα degradations in RAW264.7 macrophage cells. (A) RAW264.7 cells were pretreated with indicated concentrations of CG for 1 hr, then exposed to LPS (200 ng/ml) for 15 mins. The cell lysates were prepared and subjected to Western blotting analysis by using antibodies specific for IκB as described in the methods. (B) The relative protein levels were quantified by scanning densitometry and normalized to β-actin. The data showed that CG significantly prevented LPS-induced IκB degradations in a concentration-dependent manner. The images shown are representatives of three independent experiments that showed consistent results and the relative protein values are expressed as mean±SD for three experiments. *p<0.05 and **p<0.01 indicate statistically significant differences compared with LPS alone.
|
Akt signaling pathway mediates the anti-inflammatory activity of CG
To examine the underlying signaling pathway by which CG exerts its anti-inflammatory activity, effect of CG on LPS-stimulated phosphorylation of MAP (JNK, ERK, and p38) kinases and Akt in RAW264.7 macrophage cells was examined. Cells were pretreated with CG at indicated concentrations and then were treated with LPS (200 ng/ml) for 30 min. Data showed that no significant changes were observed in the phosphorylation levels of MAP kinases (Fig. 6). However, CG significantly resulted in the increased phosphorylation of Akt in the LPS-stimulated RAW264.7 cells in a concentration-dependent manner (Fig. 7). Furthermore, inhibition of Akt signaling pathway with wortmannin, a PI3K inhibitor, significantly abolished the CG-induced Akt phosphorylation (Fig. 8A), suggesting that activation of Akt/PI3K signaling mediate the anti-inflammatory action of CG. However, inhibition of PI3K/Akt pathway with wortmannin did not completely recover CG-induced suppression of iNOS and COX-2 expressions (Fig. 8B), suggesting that other signaling pathways rather than Akt/PI3K might be also involved in the anti-inflammatory property of CG. In addition, wortmannin also attenuated CG-mediated restoration of IκB level (Fig. 8C).
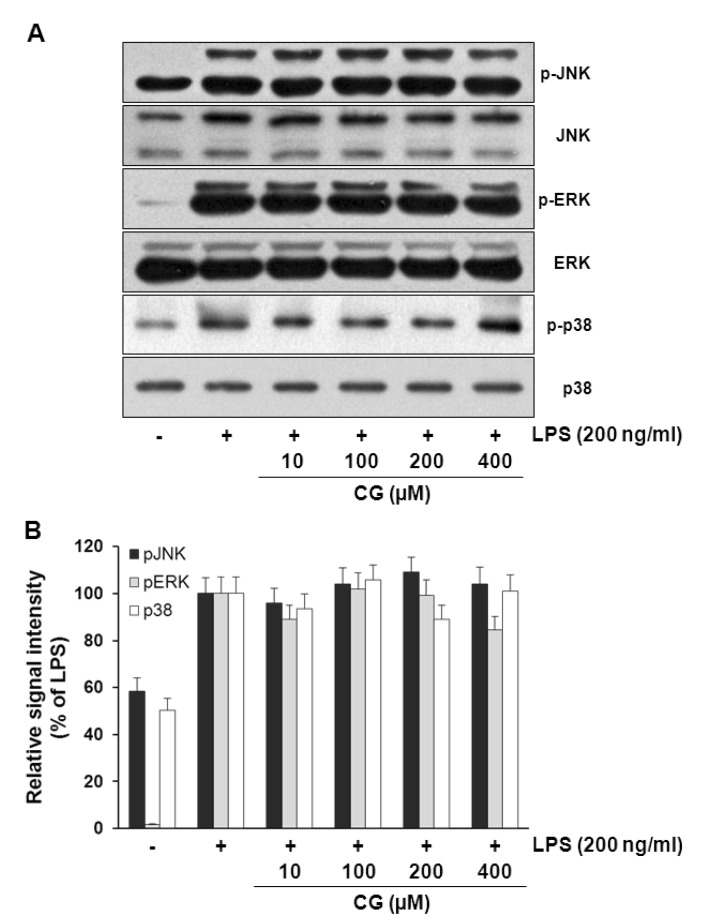 | Fig. 6CG did not affect MAPKs signaling pathways in RAW264.7 macrophage cells. (A) RAW264.7 cells were pretreated with various indicated concentrations of CG for 1 hr, then exposed to LPS (200 ng/ml) for 30 mins. The cell lysates were prepared and subjected to Western blotting analysis by using antibodies specific for total and phosphorylated forms (shown as p-) of JNK, ERK and p38. (B) The relative protein levels of p-JNK, p-ERK and p-p38 were quantified by scanning densitometry and normalized to total JNK, ERK and p38, respectively. The images shown are representatives of three independent experiments that showed consistent results and the relative protein values are expressed as mean±SD for three experiments. The data presented no effect of CG on MAPKs.
|
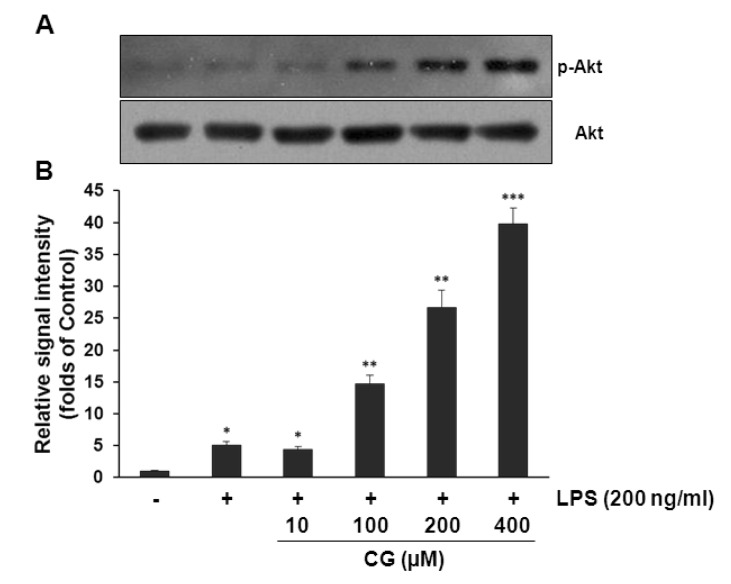 | Fig. 7CG significantly increased Akt phosphorylation in RAW264.7 macrophage cells. (A) RAW264.7 cells were pretreated with various indicated concentrations of CG for 1 hr, then exposed to LPS (200 ng/ml) for another 1 hr. The cell lysates were prepared and subjected to Western blotting analysis by using antibodies specific for total and phosphorylated forms of Akt. (B) The relative protein levels of p-Akt were quantified by scanning densitometry and normalized to total Akt. The data exhibited that CG strongly increased Akt activation in a concentration-dependent manner. The images shown are representatives of three independent experiments that showed consistent results and the relative protein values are expressed as mean±SD for three experiments. *p<0.05, **p<0.01, ***p<0.001 indicate statistically significant differences from treatments with control alone.
|
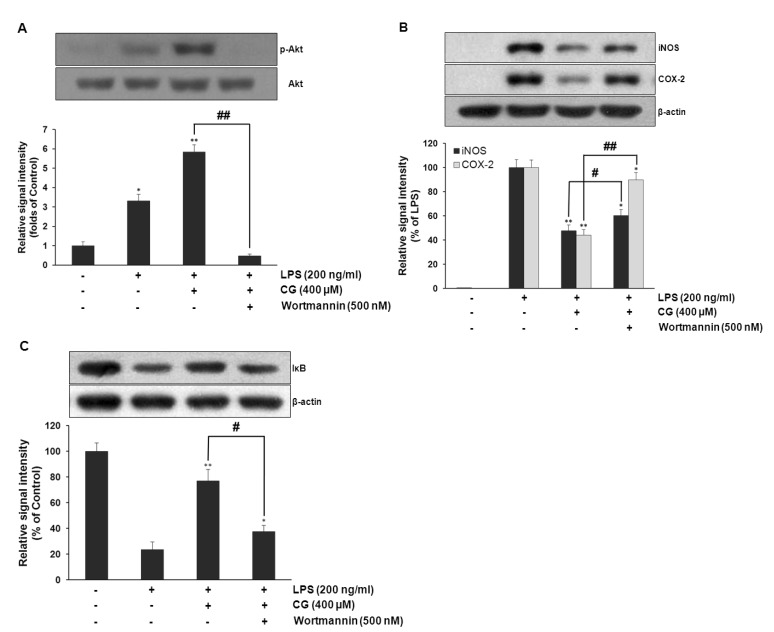 | Fig. 8PI3K/Akt was required for CG-mediated suppression of LPS-induced iNOS and COX-2 expressions and restoration of LPS-induced IκB degradation in RAW264.7 macrophage cells. RAW264.7 cells were pretreated with CG (100 µM) for 1 hr in the presence or absence of wortmannin, then exposed to LPS (200 ng/ml) for 1 hr. The cell lysates were prepared and subjected to Western blotting analysis by using antibodies specific for total and phosphorylated forms of Akt, which showed that wortmannin completely abolished CG-mediated Akt activation (A). The relative protein levels of iNOS and COX-2 were also examined. CG-mediated suppression of LPS-induced iNOS and COX-2 expressions were significantly attenuated with wortmannin but not completely (B). The protein level of IκB was examined. CG-mediated recovery of LPS-induced IκB degradation was significantly abolished with wortmannin (C). The images shown are representatives of three independent experiments that showed consistent results and the relative protein values are expressed as mean±SD for three experiments. *p<0.05, **p<0.01 indicate statistically significant differences from treatments with control alone. #p<0.05 and ##p<0.01 represent statistical significances between indicated groups.
|
Go to :
DISCUSSION
The present study clearly demonstrated that 1-p-coumaroyl β-D-glucoside (CG) exerts anti-inflammatory properties in LPS-stimulated RAW264.7 macrophage cells. CG significantly attenuated LPS-induced production of NO and PGE2 and their protein expression of iNOS and COX-2 and the secretion of pro-inflammatory cytokines such as IL-1β and TNF-α. In addition, CG showed significant attenuation of LPS-induced IκB degradation and activation of Akt phosphorylation in LPS-stimulated RAW264.7 cells.
CG is a glucose ester of p-coumaric acid, a hydroxycinnamic acid. Hydroxycinnamic acids have been reported to possess a variety of biological activities such as antioxidant, anti-tumor, anti-inflammatory properties [4,22,23]. It has been reported that p-coumaric acid suppressed cadmium-induced hepatotoxicity [24]. Recently, we observed that 3,4,5-trihydroxycinnamic acid (THC), a derivative of hydroxycinnamic acids, significantly attenuated LPS-induced inflammatory responses through the activation of a cytoprotective transcription signaling pathway of Nrf2 [6]. Ester derivatives of hydroxycinnamic acids have been demonstrated to possess numerous pharmacological activities including antitumor, antiviral, anti-inflammatory, immunosuppressive, and neuroprotective activities [1,3,7]. Previously, we reported that 1-docosanoyl cafferate (DC), a high molecular fatty alcohol ester of caffeic acid, exhibited a significant anti-inflammatory activity in LPS-stimulated murine immortalized microglial cells [25]. In addition, methyl p-hydroxycinnamate, a methyl ester derivative of p-coumaric acid, exhibited significant anti-inflammatory activity in LPS-stimulated macrophage cells (Under review). Although significant amount of coumaric acid derivatives has been identified to be present as glycosides in medicinal plants, biological activity of sugar esters of p-coumaric acid has not been extensively examined [13,26]. The present data showed that methyl p-hydroxycinnamate exhibited more potent anti-inflammatory action compared with CG. It has been also demonstrated that aglycones of flavonoid glycosides exhibit more potent free radical scavenging activity than their glycosides [27]. The present and other studies suggest that glycosides exhibit attenuated biological activities presumably with less efficient intracellular uptake due to hydrophilicity of sugar residues. Recently, we observed that avicularin, quercetin-3-α-L-arabinofuranoside, exhibited less potent anti-inflammatory property compared to its aglycone, quercetin, in LPS-stimulated macrophage cells [28]. Glycosides have been reported to be hydrolyzed to the aglycones by gycosidases in intestines and liver [29] and types of sugar moiety determine the bioavailability of glycosides [30]. Further studies are necessary to clearly elucidate the effects of a glucose residue of CG on its biological activity and efficiency of intracellular uptake.
Akt/PI3K signaling pathway has been demonstrated to exert significant anti-inflammatory effects through the suppression of NF-κB-mediated transcription [31]. The present study showed that phosphorylation of Akt was significantly increased with CG in a concentration-dependent manner in LPS-stimulated RAW264.7 cells. However, increased Akt phosphorylation was not observed with CG in naïve RAW264.7 cells, suggesting that CG-mediated Akt activation might be LPS stimulation-dependent (data not shown). Further studies are necessary to understand the stimulation-dependent manner of Akt activation by CG. Furthermore, blockade of Akt/PI3K with wortmannin, a PI3K inhibitor, significantly attenuated CG-induced Akt phosphorylation, indicating that Akt signaling pathway may play an important role in CG-induced anti-inflammatory effects. However, inhibition of PI3K/Akt pathway with wortmannin did not completely recover the CG-induced suppression of iNOS and COX-2 expression, suggesting that although Akt might play a major role in CG-mediated anti-inflammatory actions, other signaling pathways might be also involved in the anti-inflammatory property of CG. MAPK signaling pathways have been reported to play certain roles in anti-inflammatory mechanism [32]. However, no noticeable changes of MAPKs were not observed in the present study, suggesting that the anti-inflammatory activity of CG presumably be through the activation of Akt, but not MAPK, signaling pathway in the present cell culture model. However, further studies are necessary to delineate the exact mechanism by which CG activates Akt and other signaling pathways.
Macrophages and monocytes play essential roles in the host defense against bacterial infection [15]. However, aberrantly activated macrophages also play a detrimental role in inflammation-related disorders including sepsis by producing various pro-inflammatory mediators and cytokines [18]. Macrophages initiate LPS-induced pro-inflammatory gene transcription when LPS binds to its membrane receptor, TLR4, which leads to the phosphorylation of multiple kinases, which subsequently activates various transcription factors such as NF-κB and AP-1 families [33,34]. We previously reported that LPS causes increased expression of pro-inflammatory mediators and the increased degradation IκB [28,35]. CG significantly attenuated LPS-induced IκB degradation in a concentration-dependent manner. In addition, CG significantly attenuated LPS-induced extracellular secretion of pro-inflammatory cytokines such as IL-1β and TNF-α. Although pro-inflammatory cytokines such as IL-1β and TNF-α, and NF-κ B-mediated transcription have been reported to play key roles in inflammatory responses, other cytokines and transcription factors might also play a certain role. Therefore, further studies are necessary to clearly explain the effect of CG other cytokines and transcription factors in LPS-mediated inflammatory conditions.
NF-κB is a key transcription factor for the expression of a variety of pro-inflammatory genes such as iNOS, COX-2, and cytokines [36,37]. The aberrant activation of NF-κB has been widely demonstrated to be associated with various pathological conditions including cancers and autoimmune diseases [38,39]. In endotoxin-stimulated macrophages, NF-κB translocates to the nucleus for the transcription of pro-inflammatory genes when IκB is phosphorylated with LPS-induced IκB Kinase (IKK) and subsequently degraded through proteasomal pathway [40,41]. The present data showed that CG significantly attenuated LPS-induced IκB degradation in a concentration-dependent manner, which is suggestive of subsequent prevention of nuclear translocation of NF-κB. We previously observed that inhibition of IκB degradation eventually retains p65 subunit of NF-κB in cytoplasm [35].
In conclusion, the present study clearly demonstrates that CG possesses anti-inflammatory property such as suppression of NO and PGE2 production and cytokine secretion via the inhibition of nuclear translocation of NF-κB in LPS-stimulated RAW264.7 macrophage cells. The anti-inflammatory action of CG is considered to be mediated through the activation of Akt/PI3K signaling pathway. However, further studies are necessary to delineate the exact mechanism by which CG activates Akt/PI3K signaling pathway. The present study strongly suggests that CG might be a valuable therapeutic agent in the treatment of inflammatory pathogenic conditions including sepsis.
Go to :
 XML Download
XML Download