

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Chronic stress contributes to the development of depression-like symptoms, mood and anxiety disorders [1,2]. It is well known that learning and memory is affected by stress [3,4] and that prolonged exposure to stress can affect the region of the brain that is central to learning and emotional responses: the hippocampus, amygdala and prefrontal cortex [1,2,5].
Dehydroevodiamine·HCl (DHED), a component separated from Evodia rutaecarpa Bentham, improves memory deficit of Alzheimer's mice models such as Tg2576 and APP-CT Tg mice and the scopolamine induced amnesia model [6-8].
The main purpose of this study was to investigate the effects of DHED on stress. We examined whether DHED could improve memory impairments and depression-like behaviors. We checked the effects of DHED or fluoxetine, a well-known antidepressant on stress-induced memory impairments and depression-like behaviors, using open-field test, Y-maze performance and forced swimming test.
In addition, we examined whether DHED could elevate stress-induced the low levels of neural cell adhesion molecule (NCAM). Chronic stress induced cognitive and structural alterations were also shown to reduce the expression of NCAM [9-11]. NCAM is known to link to the development of neuronal damage under stress condition [12].
In our study, we provide in vitro and in vivo evidences that DHED can improve stress-induced depression and memory impairment and significantly recover the low levels of NCAM proteins caused by stress.
METHODS
Experimental protocol
Five weeks-old male Wistar rats weighing 160~180 g were used. Immobilization was used to induce chronic stress according to the previously described method [8,13,14]. Fluoxetine·HCl (10 mg/kg/day, p.o.) or DHED (10 mg/kg/day, p.o., Jeil Pharm., Korea) dissolved in 0.3% CMC (carboxymethylcellulose, Sigma, USA) solution was administered via oral route to animals according to the treatment schedule in Fig. 1A (n=4~5 per group). All behavior tests were scored by an investigator blinded to treatment conditions and all manipulations were performed in accordance with the Guidelines for Animal Experiments of Ethics Committee of Seoul National University.
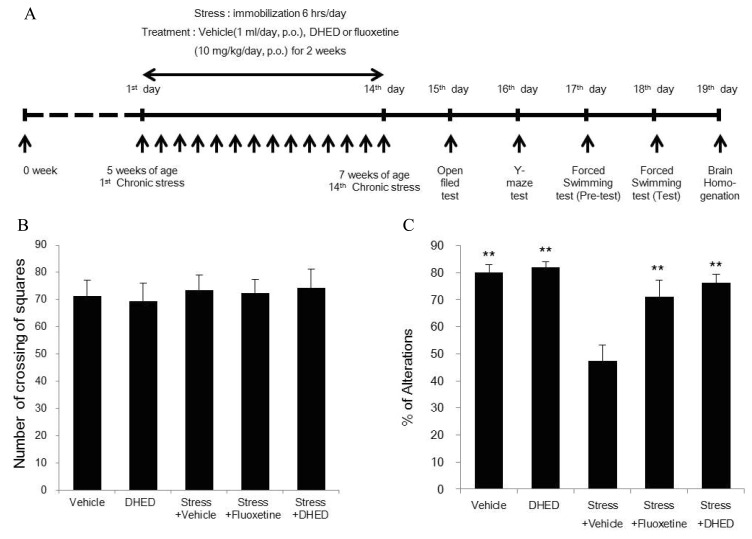
Fig. 1
Effects of DHED on locomotor activity and memory impairment induced by stress in Open field test (B), Y-maze test (C). (A) Behavior tests were performed following repeated administrations of vehicle, DHED or fluoxetine for 14 days. (B) In open field test, there was no significant difference in the number of crossings compared with the control group. (C) In Y-maze test, percentage of success was significantly increased by DHED or fluoxetine treatment compared with the vehicle-treated stressed rats. Data represents mean±SEM. ANOVA followed by the LSD test (n=4~5, **p<0.01, *p<0.05).
![]()
Behavior test
Locomotion was measured after stress. Rats were freely placed in the center of the 40×50×60 cm open filed divided into 30 equal squares by black lines. The numbers of crossings of the black lines were used as measures of locomotion for 5 min [10]. Y-maze test was used for testing spatial working memory during the 8-min testing session according to the previously described method [10]. The apparatus was an opaque plastic maze and each arm was 80 cm long, 35 cm high, and 15 cm wide, and converged at an equilateral angle. Food and water were restricted. Three continuous entries into three different arms were counted as an alternation. Forced swimming test has been widely used to evaluate potential effectiveness of antidepressant drugs according to the previously described method [15]. The rats were gently placed individually in plexiglass cylinders (40 cm height; 18 cm in diameter, 30 cm water depth), and contained water was at a temperature of 25℃, and then the total duration of swimming mobility (vigorous attempts to escape) time was measured for a 10 min test session.
Western blotting
After perfusion, rat brains were lysed in RIPA buffer with cocktail of protease inhibitors (Roche). The proteins were separated by SDS-PAGE and transferred to a PVDF membrane as described [14]. The blot was probed with NCAM (Millipore, USA, AB5032) and GAPDH (Santa Cruz Biotechnology, USA, SC-257788) antibodies. After being incubated with horseradish peroxidase-conjugated secondary antibody, the membrane was visualized by ECL plus solution (Amersham Pharmacia).
Preparation of primary cortical neurons
Primary neurons were derived from the embryos (E17 days) cortex of 6- to 7-week-old pregnant SD rat (Koatech, South Korea). Briefly, an anesthetized pregnant rat was exposed to an overdose inhalation of 70~100% CO2 gas for euthanasia, and then the uterus was extracted into cold phosphate buffered saline (PBS). We collected embryos in cold PBS and then moved them into a filtered flask to administer euthanasia using an overdose inhalation of 70~100 % CO2 gas. Next, we placed them in a fresh dish with HBSS. The cerebral cortex was dissected from embryo rat brains and dissociated by gently triturate. Using curved, sharp forceps, the brains were collected from the embryos under a microscope. All animal experimental procedures were performed in accordance with 'the Guidelines of the Ethics Committee at Seoul National University'.
Cells were cultured in a specified medium for neurons (neurobasal medium supplemented with B27 [Gibco BRL, NY] and penicillin-streptomycin (100 U/ml/100 mg/ml) [Life Technology] at 37℃ and 5% CO2 and plated at a density of 5×104 cells/well on Deckgläser cover glasses (18 mm Φ, marienfeld) coated with PLL (1 mg/ml) in the 12 well plates.
Experiments were performed on 10-day cultures. After DHED (0.5 and 1 µM) and fluoxetine (0.5 µM) treatment for 4 hr, staurosporin (1 µM) were added for 1 hr.
Immunocytochemistry
After three washes with PBS, cells were fixed with 4% p-formaldehyde for 30 min. After three washes with PBS, cells were blocked and permeabilized in PBS added with 2% FBS, 2% BSA, 4% normal goat serum and 0.2% Triton X-100. Cells were incubated with primary antibodies for overnight at 4℃. After washing with PBS, proteins were visualized using secondary species-specific IgG conjugated with fluorescence. Cells were observed by confocal laser-scanning microscopy using an LSM-510 imaging system (Carl Zeiss Inc., Germany).
RESULTS
DHED improved stress-induced memory deficits and stress-induced mobility reduction in behavioral tests
We examined the effects of DHED and fluoxetine on locomotor activity in open field test. There were no noticeable differences in the numbers of crossings among five groups, indicating that locomotor activity is not affected by DHED or fluoxetine (Fig. 1B). Then, we investigated whether stress-induced spatial memory deficits could be improved by the treatment with DHED or fluoxetine in Y-maze test. As shown in Fig. 1C, the percentage of alternation behavior of the stressed rats (47.37±5.85) was more decreased than that of the vehicle-treated control (86.67±2.80) or the DHED-treated (82.03±1.88) control. But, the stress-induced decline in spontaneous alternations was significantly elevated to the vehicle control level by DHED (76.33±3.04) or fluoxetine treatment (71.08±6.18). Forced swimming test was done [15] to determine whether stress-induced reduction in the mobility time could be changed by the treatment with DHED or fluoxetine. As shown in Fig. 2, the climbing mobility time of the vehicle-treated stressed rats (136.0±13.71) was decreased in comparison to that of the vehicle-treated (200.5±21.20) or DHED-treated control (236.0±26.88). But, the stress-induced reduction in the mobility time was significantly increased by DHED (249.75±17.56) or fluoxetine treatment (262.8±31.39).
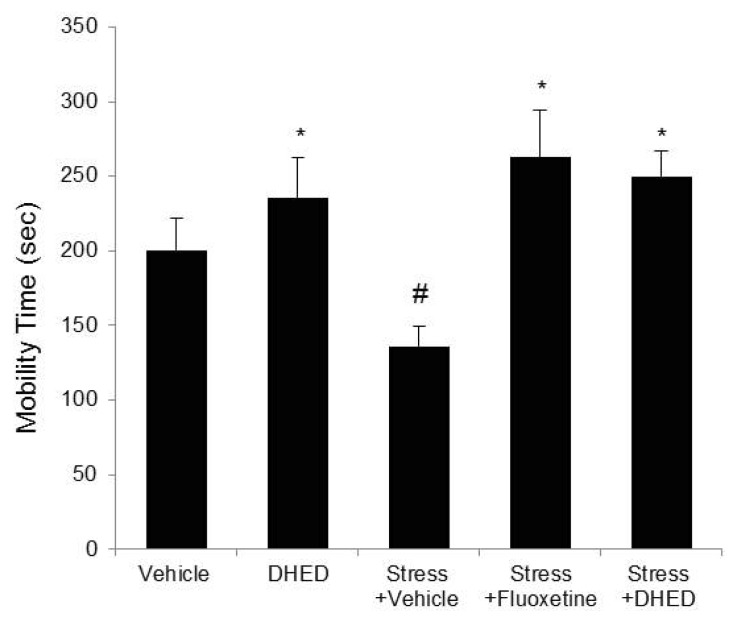
Fig. 2
Effects of DHED on depression like behavior induced by stress in Forced swimming test. On forced swimming test, mobility time was increased significantly in DHED or fluoxetine treated stressed rats compared with the vehicle-treated stressed rats. Data represents mean±SEM. ANOVA followed by the LSD test (n=4~5, **p<0.01, *p<0.05).
![]()
DHED prevented the stress-induced decrease in total NCAM levels of rat brains
To assess DHED's therapeutic effects, we examined the levels of NCAM (180, 140, or 120 kDa isoform) that involved with synaptic plasticity and development of neuron [12] in the brain total homogenates. NCAM was detected by western blot with a specific antibody against NCAM and quantified (Fig. 3A). The levels of total NCAM in the vehicle-treated stressed rats were decreased in comparison to those of the vehicle- (1.588±0.202) or DHED-treated (1.995±0.228) control rats (Fig. 3B). However, the reduction of total NCAM levels (0.714±0.096) induced by stress was prevented by DHED (1.879±0.274) or fluoxetine (1.036±0.191) treatment.

Fig. 3
DHED rescues the low NCAM levels by stress. (A) Representative immune-blot for total NCAM in the rat brains of the vehicle- and DHED-treated and Vehicle-, fluoxetine- or DHED-treated stressed rat were shown. Total NCAM levels in brain homogenates were examined after the behavioral tests by Western blotting. (B) The relative quantity of total NCAM levels were significantly increased by DHED treatment compared with the vehicle-treated stressed rats (n=4, *p<0.05 by ANOVA followed by the LSD test).
![]()
To further confirm the preventive effects of DHED or fluoxetine in the level of NCAM reduced by stress, we checked the NCAM levels in primary cortical neurons treated with staurosporine which is a prototypical ATP-competitive kinase inhibitor in that it binds to many kinases with high affinity.
NCAM levels were also decreased by the treatment with staurosporine (1 µM) in primary cortical neurons (Supplementary Fig 1). The pretreatment with DHED (0.5 and 1 µM) or fluoxetine (0.5 µM) restored the level of NCAM reduced by sturosporine.
DISCUSSION
The present study strongly suggests that DHED improved memory impairments and depression-like behaviors induced by stress in rats.
Firstly, we found that both DHED and fluoxetine could alleviate memory impairments induced by stress because DHED improved cognitive impairments of other amnesic animal models in previous studies [7]. Fluoxetine is widely used for a spectrum of mood disorders [16] and also protects against the adverse effects of different types of stressors [17]. Several studies demonstrated that fluoxetine is the treatment of choice for depression due to a better safety profile and improved tolerability compared to the older tricyclic antidepressants [16]. Here, we used fluoxetine as a positive control. Stress can induce cognitive impairment as well as depression-like behaviors. In rats, chronic stress was reported to induce spatial memory impairments [1]. Especially, the reduction of alteration behaviors in Y-maze test shows the impairment of memory [10]. The present results demonstrate that the stress-induced decrease in the percentage of success was elevated to the control level by DHED treatment by fluoxtine treatment.
Secondly, we also found that DHED or fluoxetine could improve depressive behavior in rats. Chronic stress plays a pivotal role in several neuropsychiatric disorders such as depression [18]. A depressive behavior is related to the increase of immobility time [15,19] and antidepressants decrease immobility in forced swimming test. In this study, the immobility time of stress group was obviously higher than that of the control group, but it was reduced to the control level by DHED or fluoxetine. Our results show that DHED could improve depression-like behaviors as well as memory impairments induced by stress without causing change in mobility.
It is well known that NCAM is expressed on the membrane of neurons and glial cells and involves plasticity and development of neuron [12,20]. Chronic restraint stress in adult rats decreases NCAM mRNA and protein expression in the hippocampus and other brain regions [9,20]. Our results indicate that total NCAM levels of the vehicle-treated stressed rats were decreased than those of the vehicle control. But it was significantly recovered to the vehicle control level by DHED or fluoxetine treatment. The expression of each NCAM isoform were reported to be variable in each group, therefore many researchers prefer the comparison of the concentration of total NCAM to that of each NCAM isoform [9,20]. Therefore, our results suggest that DHED might increase brain activity and memory partially through increasing total NCAM levels which are related to neuronal plasticity and regeneration. Additionally, we tested whether DHED can affect intracellular NCAM levels in neurons. Interestingly, DHED prevented the decrease in NCAM levels by staurosporine. It has been reported that staurosporine induces apoptosis [21] and that NCAM is linked to the development of neuronal damage [12].
In conclusion, DHED has beneficial effects on memory impairments, depression-like behaviors, decreased neuronal viability and NCAM levels induced by stress. Therefore, DHED as well as fluoxetine may be a potential drug against stress-induced memory impairments and depression-like behaviors.
 XML Download
XML Download