

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Previously, Lim and his coworkers [1,2,3] have reported that all of total giseng saponin (TGS), panaxadiol and panaxatriol cause the increased secretion of catecholamines (CA) from the isolated perfused rabbit adrenal glands in a Ca2+-dependent fashion, which are mediated by the activation of cholinergic (both nicotinic and muscarinic) receptors and partly the direct action on the rabbit adrenomedullary chromaffin cells. However, it has been reported that TGS shows the inhibitory effect on the CA release evoked by stimulation of nicotinic receptors in the perfused rat adrenal medulla [4], and also that several ginseng saponins (ginsenosides) from the ginseng root inhibit the CA secretion from bovine adrenal chromaffin cells stimulated by ACh [5,6,7,8].
In previous study, Tachikawa and his co-workers [6] have shown that most of the ginsenosides (1~100 µM) had a tendency to reduce the ACh-evoked CA secretion. The order of inhibitory potency (at the concentration of 10 µM) was as follows: Rg2>Rf>Re>Rh1>Rb2, Rg1>Rb1>Rc>Rb3, Rd, Ro, Rs1. The inhibition of ginsenoside-Rg2 at 10 µM was 72%, but ginsenosides Rb3, Rd, Ro and Rs1 did not show the inhibitory effect.
In the conscious rats, the releasing effect of NO of Korean Red Ginseng (KRG) may be partly contributed to the hypotensive effect [9]. It has also been shown that Rg3 relaxes the rat thoracic aorta as a consequence of NO production [10]. Han and his co-workers [11] have found evidence that NO levels in exhaled breath of human volunteers by KRG were increased along with decreased blood pressure and heart rate. In a series of studies, it has been found that ginsenosides reduce blood pressure via increases in production of endothelial nitric oxide [12] and that Rg3 is the most potent ginsenoside that activates eNOS in rat aorta [13]. Although some investigators have demonstrated that Rg3 induces eNOS activation in the vasculature of animal models [12,14], Hien and his co-workers [15] have found that Rg3 activates eNOS via eNOS phosphorylation in ECV 304 human endothelial cells and increases in eNOS expression.
Several studies have shown that ginsenoside Rg1 can cause endothelial-dependent relaxation in the rat aorta [14] and enhance endogenous NO production in human umbilical vein endothelial cells [16], rat kidney [17] and in porcine coronary arteries [18].
Despite of these many studies on various ginseng saponins, there are still a little known effects of ginsenoside-Rb2 on adrenal CA secretion. Therefore, in this study, we investigated whether ginsenoside-Rb2 affects nicotinic receptor stimulation-evoked CA secretion from the perfused model of the isolated rat adrenal medulla and tried to establish its mechanism of action.
METHODS
Experimental procedure
Mature male Sprague-Dowley rats (purchased from DAMOOL SCIENCE, International Customer Service, Seoul, Korea), weighing 200 to 300 grams, were used in the experiment. The animals were housed individually in separate cages, and food (Cheil Animal Chow) and tap water were allowed ad libitum for at least a week to adapt to experimental circumstances. On the day of experiment, a rat was anesthetized with thiopental sodium (50 mg/kg) intraperitoneally, and tied in supine position on fixing panel.
1. Isolation of adrenal glands
The adrenal gland was isolated by the modification of previous method [19]. The abdomen was opened by a midline incision, and the left adrenal gland and surrounding area were exposed by the placement of three-hook retractors. The stomach, intestine and portion of the liver were not removed, but pushed over to the right side and covered by saline-soaked gauge pads and urine in bladder was removed in order to obtain enough working space for tying blood vessels and cannulations. A cannula, used for perfusion of the adrenal gland, was inserted into the distal end of the renal vein after all branches of adrenal vein (if any), vena cava and aorta were ligated. Heparin (400 IU/mL) was injected into vena cava to prevent blood coagulation before ligating vessels and cannulations. A small slit was made into the adrenal cortex just opposite entrance of adrenal vein. Perfusion of the gland was started, making sure that no leakage was present, and the perfusion fluid escaped only from the slit made in adrenal cortex. Then the adrenal gland, along with ligated blood vessels and the cannula, was carefully removed from the animal and placed on a platform of a leucite chamber. The chamber was continuously circulated with water heated at 37±1℃ (Fig. 1).
Perfusion of adrenal gland
The adrenal glands were perfused by means of peristaltic pump (Isco, St. Lincoln, NE, USA) at a rate of 0.31 mL/min. The perfusion was carried out with Krebs-bicarbonate solution of following composition (mM): NaCl, 118.4; KCl, 4.7; CaCl2, 2.5; MgCl2, 1.18; NaHCO3, 25; KH2PO4, 1.2; glucose, 11.7. The solution was constantly bubbled with 95 % O2+5% CO2 and the final pH of the solution was maintained at 7.4~7.5. The solution contained disodium EDTA (10 µg/mL) and ascorbic acid (100 µg/mL) to prevent oxidation of catecholamines.
Drug administration
The perfusions of DMPP (10-4 M) for 2 minutes and/or a single injection of ACh (5.32×10-3 M) and KCl (5.6×10-2 M) in a volume of 0.05 mL were made into perfusion stream via a three-way stopcock, respectively. McN-A-343 (10-4 M), veratridine (10-4 M), Bay-K-8644 (10-5 M) and cyclopiazonic acid (10-5 M) were also perfused for 4 min, respectively.
In the preliminary experiments, it was found that upon administration of the above drugs, secretory responses to ACh, KCl, McN-A-343, Bay-K-8644 and cyclopiazonic acid returned to preinjection level in about 4 min, but the responses to DMPP in 8 min.
Collection of perfusate
As a rule, prior to stimulation with various secretagogues, the perfusate was collected for 4 min to determine the spontaneous secretion of CA (background sample). Immediately after the collection of the background sample, collection of the perfusates was continued in another tube as soon as the perfusion medium containing the stimulatory agent reached the adrenal gland. Stimulated sample's perfusate was collected for 4 to 8 min. The amounts secreted in the background sample have been subtracted from that secreted from the stimulated sample to obtain the net secretion value of CA, which is shown in all of the figures.
To study the effect of Rb2 on the spontaneous and evoked secretion, the adrenal gland was perfused with normal Krebs solution for 90 min, and then the perfusate was collected for a certain period (background sample). Then the medium was changed to the one containing the stimulating agent or along with Rb2, and the perfusates were collected for the same period as that for the background sample. The adrenal gland's perfusate was collected in chilled tubes.
Measurement of catecholamines
The content of CA (all of epinephrine, norepinephrine and dopamine) in perfusate was measured directly by the fluorometric method of Anton and Sayre [20] without the intermediate purification alumina for the reasons described earlier [19] using fluorospectrophotometer (Kontron Co., Milano, Italy).
A volume of 0.2 mL of the perfusate was used for the reaction. The CA content in the perfusate of stimulated glands by secretagogues used in the present work was high enough to obtain readings several folds greater than the reading of control samples (unstimulated). The sample blanks were also lowest for perfusates of stimulated and non-stimulated samples. The CA content in the perfusate was expressed in terms of norepinephrine (base) equivalents.
Measurement of NO release
NO release was measured using a NO-selective microelectrode (ami700, Innovative Instruments Inc) and an amplifier (inNo meter, Innovative Instruments Inc). Adrenomedullary NO production was quantified as the integrated signal detected by the microelectrode after perfusion of Rb2 into rat adrenal medulla, as previously described [21]. The electrode was calibrated by producing standardized concentrations of NO in 0.5% (wt/vol) KI in 0.1 Mol/L H2SO4 from NaNO2 standards. NO release was quantitated as the current detected at the electrode after loading Rb2 into adrenal medulla. NO release was calculated as picomoles.
Statistical analysis
The statistical difference between the control and the pretreated groups was determined by the Student's t and ANOVA tests. A p-value of less than 0.05 was considered to represent statistically significant changes unless specifically noted in the text. Values given in the text refer to means and the standard errors of the mean (S.E.M.). The statistical analysis of the experimental results was made by computer program described by Tallarida and Murray [22].
Drugs and their sources
The following drugs were used: 1.1-dimethyl-4-phenyl piperazinium iodide (DMPP), acetylcholine chloride, norepinephrine bitartrate, potassium chloride (KCl), Nω-nitro-L-arginine methyl ester hydrochloride (L-NAME), methyl-1,4-dihydro-2,6-dimethyl-3-nitro-4-(2-trifluoro-methylphenyl)-pyridine-5-carboxylate (BAY-K-8644), cyclopiazonic acid, veratridine and potassium nitrate (Sigma Chemical Co., U.S.A.). Ginsenoside-Rb2 was a gift from Dr. Geon-Wook Kang (Professor of Seoul National University, Seoul, Korea). Drugs were dissolved in distilled water (stock) and added to the normal Krebs solution as required except Bay-K-8644, which was dissolved in 99.5% ethanol and diluted appropriately with Krebs-bicarbonate solution (final concentration of alcohol was less than 0.1%). Concentrations of all drugs were expressed in terms of molar base.
RESULTS
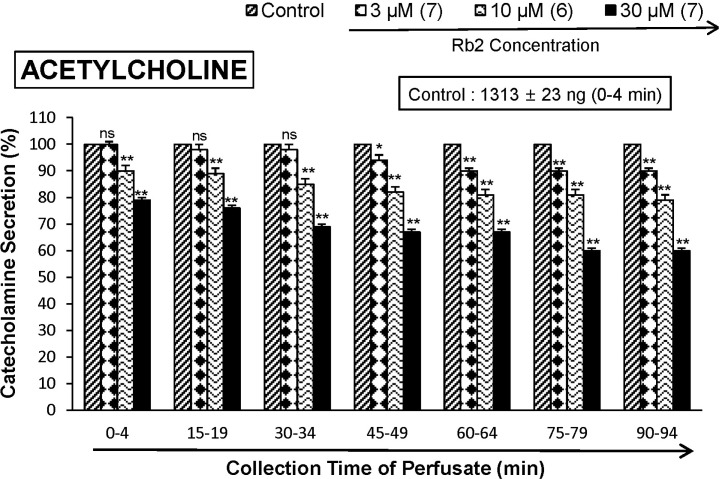
Influence of Ginsenoside-Rb2 on the CA secretion evoked by ACh, high K+, and DMPP from the perfused rat adrenal medulla
After the perfusion with oxygenated Krebs-bicarbonate solution for 1 hr, basal CA release from the isolated perfused rat adrenal glands amounted to 20±2 ng for 2 min (n=12). Since Tachikawa and his co-workers [6] have reported that most of the ginsenosides (1~100 µM) including Rb2 had a tendency to reduce the ACh-evoked CA secretion, it was attempted initially to examine the effects of Rb2 itself on CA secretion from the perfused model of the rat adrenal glands. However, in the present study, Rb2 itself did not produce any effect on basal CA output from the perfused rat adrenal glands (data not shown). Therefore, it was decided to investigate the effects of Rb2 on cholinergic nicotinic receptor stimulation- as well as membrane depolarization-evoked CA secretory responses. Secretagogues were given at 15 to 20 min-intervals. Rb2 was present for 90 minutes after the establishment of the control release.
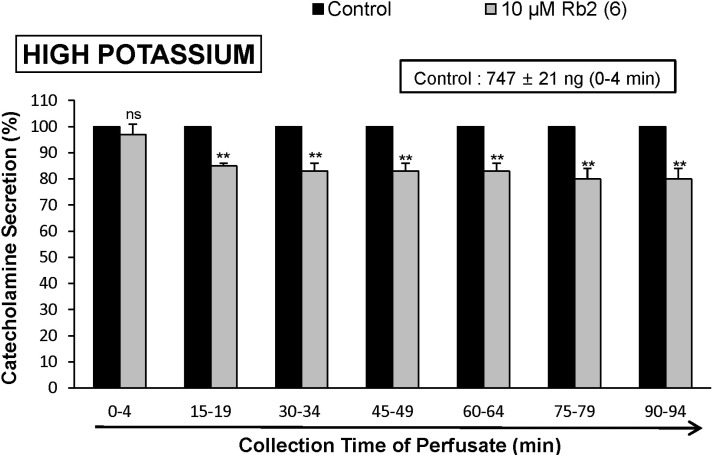
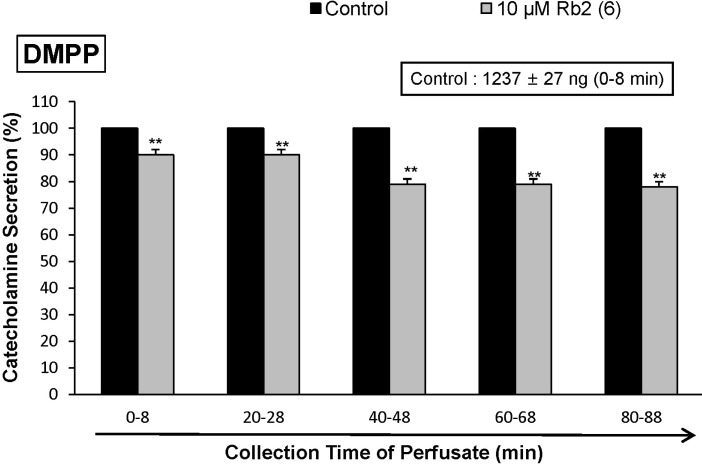
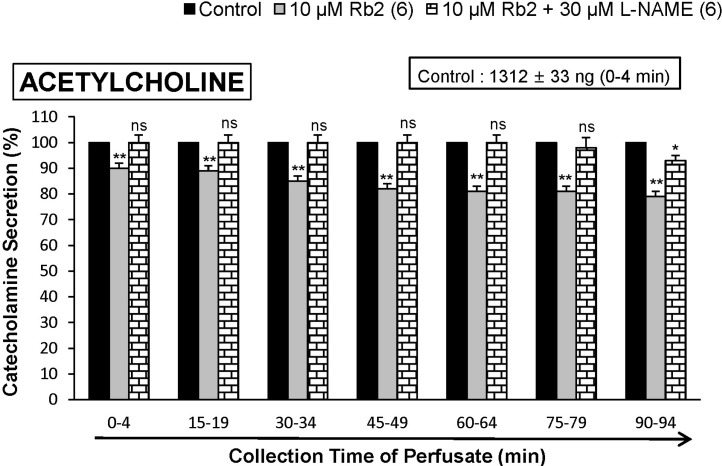
When ACh (5.32×10-3 M) in a volume of 0.05 mL was injected into the perfusion stream, the amount of CA secreted was 1313±23 ng for 4 min. However, the pretreatment with Rb2 in the range of 10~30 µM for 90 min relatively concentration- and time-dependently inhibited AChstimulated CA secretion. As shown in Fig. 1, in the presence of Rb2, the CA releasing responses were inhibited to 60% of the corresponding control release (100%). Also, it has been shown that depolarizing agent like high KCl stimulates markedly CA secretion (747±21 ng for 0~4 min). High K+ (5.6×10-2 M)-stimulated CA secretion after the pretreatment with 10 µM Rb2 was maximally inhibited to 80% of the control after 75 min period, although it was not initially affected for the first period (0-4 min) as shown in Fig. 2. DMPP (10-4 M), which is a selective nicotinic receptor agonist in autonomic sympathetic ganglia, evoked a sharp and rapid increase in CA secretion (1237±27 ng for 0~8 min). However, as shown in Fig. 3, DMPP-stimulated CA secretion after the pretreatment with Rb2 (10 µM) was greatly reduced to 78% of the control release.
Influence of Ginsenoside-Rb2 on the CA secretion evoked by Bay-K-8644, cyclopiazonic acid and veratridine from the perfused rat adrenal medulla
Since Bay-K-8644 is known to be a calcium channel activator, which enhances basal Ca2+ uptake [23] and CA release [24], it was of interest to determine the effects of Rb2 on Bay-K-8644-stimulated CA secretion from the isolated perfused rat adrenal glands. Bay-K-8644 (10-5 M)-stimulated CA secretion in the presence of Rb2 (10 µM) was reduced to 73% of the control except for the early 15 min period as compared to the corresponding control release (640±33 ng for 0~4 min) from 6 rat adrenal glands, as shown in Fig. 4.
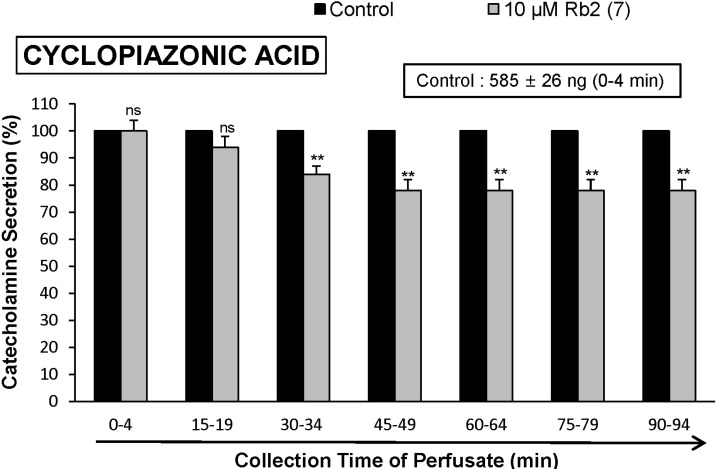
Cyclopiazonic acid, a mycotoxin from Aspergillus and Penicillium, has been described as a highly selective inhibitor of Ca2+-ATPase in skeletal muscle sarcoplasmic reticulum [25,26]. The inhibitory action of Rb2 on cyclopiazonic acid-evoked CA secretory response was observed as shown in Fig. 5. In the presence of Rb2 (10 µM) in 7 rat adrenal glands, cyclopiazonic acid (10-5 M)-evoked CA secretion was also inhibited to 78% of the control response (585±26 ng for 0~4 min), although it was not initially affected for the first and second periods (0~19 min).
It has been known that veratridine-induced Na+ influx mediated through voltage-dependent Na+ channels increased Ca2+ influx via activation of voltage-dependent Ca2+ channels and produced the exocytotic CA secretion in cultured bovine adrenal medullary cells [27]. As shown in Fig. 6, veratridine (5×10-5 M) sharply produced CA secretion (848±23 ng for 0~4 min). In 8 rat adrenal glands, Rb2 (10 µM) also attenuated veratridine-induced CA secretion to 74% of the corresponding control release.
Influence of Ginsenoside-Rb2 plus L-NAME on Ach-Evoked CA secretion from the perfused rat adrenal medulla
It has also been found that, in this study, Rb2 inhibited the CA secretory response evoked by stimulation of cholinergic receptors in the perfused rat adrenal glands. Therefore, to study the relationship between NO and Rb2-induced inhibitory effects on the CA release from the rat adrenal glands, the effect of L-NAME on Rb2-induced inhibitory responses of CA secretion evoked by cholinergic receptorstimulation as well as membrane depolarization was examined. In the present study, in the simultaneous presence of Rb2 (10 µM) and L-NAME (30 µM) for 90 min from 6 rat adrenal glands, ACh (5.32×10-3 M)-evoked CA release was almostly recovered to 100~93% of the corresponding control level compared to that of Rb2 (10 µM)-treated alone, as ishown in Fig. 7.
Influence of Ginsenoside-Rb2on the level of nitric oxide released from the perfused rat adrenal medulla
As shown in Fig. 7, the inhibitory effects of Rb2 on AChevoked CA secretory responses were significantly recovered to the control level in the presence of L-NAME. Therefore, it was decided directly to determine the level of NO released from rat adrenal medulla after the treatment of Rb2. Moreover, it has been found that ginsenosides reduce blood pressure via increases in production of endothelial nitric oxide [12] and that ginsenoside Rg3 is the most potent ginsenoside that activates eNOS in rat aorta [13]. Although some investigators have demonstrated that Rg3 induces eNOS activation in the vasculature of animal models [13,14].
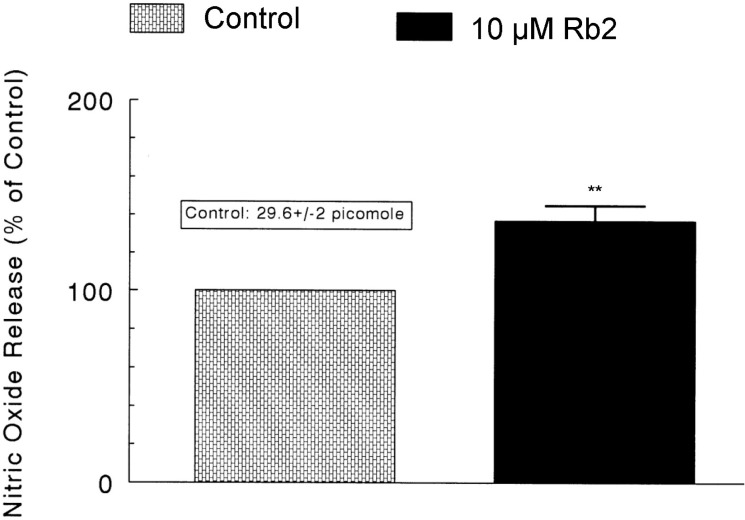
In 6 adrenal glands, the basal amount of NO released from medulla prior to administration of Rb2 was 29.6±2 picomoles. However, 8 min after loading with Rb2 (10 µM) it was greatly elevated to 40.6±2.5 picomoles, which was 137% of the basal release, as shown in Fig. 8.
DISCUSSION
The present experimental results showed that Rb2 inhibited the CA secretory responses from the isolated perfused rat adrenal gland evoked by stimulation of cholinergic nicotinic receptors as well as by direct membrane-depolarization. It seems that this inhibitory effect of Rb2 is exerted by inhibiting influx of both Na+ and Ca2+ ions through their channels into the adrenomedullary chromaffin cells and also by suppression of Ca2+ release from the cytoplasmic calcium store at least partly via increased NO production due to activation of neuronal NO synthase, which is relevant to the blockade of neuronal nicotinic receptors.
In the present study, in the simultaneous presence of Rb2 and L-NAME (an NO synthase inhibitor), the ACh-evoked CA secretory responses was considerably recovered to the extent of the corresponding control secretion compared to those of Rb2-treatment alone. This result is well consistent with the report that, in a series of studies, ginsenosides reduce blood pressure via increases in production of endothelial nitric oxide [12] and that ginsenoside Rg3 is the most potent ginsenoside that activates eNOS in rat aorta [13]. Although several investigators have demonstrated that Rg3 induces eNOS activation in the vasculature of animal models [12,14], Hien and his co-workers [15] have found that Rg3 activates eNOS via eNOS phosphorylation in ECV 304 human endothelial cells and increases in eNOS expression. Moreover, in this study, following treatment of Rb2 into adrenal medulla, NO production was greatly elevated as shown in Fig. 8. Taking into account these findings, in the present study, it is likely that Rb2 inhibits the CA secretory response evoked by various secretagogues through increasing NO production in adrenal chromaffin cells, since in the simultaneous presence of Rb2 and L-NAME, an inhibitor of NO synthase, ACh-evoked-CA secretion was nearly recovered to the corresponding control level compared to that of Rb2-treatment alone, and also Rb2 notably enhanced NO release from rat adrenal medulla.
In the support of this finding, it has been reported that the NOS inhibitor, L-NAME enhances K+-stimulated CA secretion in cultured bovine chromaffin cells [28] and that sodium nitroprusside (SNP) inhibits ACh-induced CA secretion in bovine chromaffin cells [29]. These studies suggest that NO may play an inhibitory role in the control of the CA secretion. Moreover, the presence of endothelial cells has been reported to inhibit the K+-induced or the nicotinic receptor agonist DMPP-induced CA secretion in cultured bovine chromaffin cells [28], suggesting that not merely nNOS but also eNOS may play roles in modulating adrenal CA secretion. In view of the results so far reported, the present studies strongly suggest that Rb2 possesses the ability partly to activate nNOS in the adrenomedullary chromaffin cells, resulting in enhancement of NO as well as inhibition of the CA release, in addition to the direct inhibitory effects on the CA secretion. In supporting this finding, among the ginsenosides of the protopanaxatriol and protopanaxadiol groups, ginsenoside Rg3 is the most potent vasodilator. Previously, it was shown that Rg3 inhibited calcium-induced vascular contraction [13] as well as phenylephrine-induced vasocontraction as a consequence of NO production [10].
In contrast, it has been reported that L-NAME inhibits acetylcholine (ACh)-induced CA secretion in bovine chromaffin cells [29], and that the NO donor SNP enhances nicotine-induced CA secretion in cultured bovine chromaffin cells [30]. These findings suggest that NO may facilitate cholinergic agonist-induced CA secretion. On the other hand, a few in vivo studies have suggested that NO does not play a role in regulation of adrenal CA secretion [31,32].
Generally, the adrenal medulla has been employed as a model system to study numerous cellular functions involving not only noradrenergic nerve cells but also neurons. During neurogenic stimulation of the adrenal medulla, ACh is released from splanchnic nerve endings and activates cholinergic receptors on the chromaffin cell membrane [33]. This activation initiates a series of events known as stimulus-secretion coupling, culminating in the exocytotic release of CA and other components of the secretory vesicles into the extracellular space. Usually, two mechanisms are involved in the secretion of adrenal medullary hormones. Upon excitation of splanchnic nerves, ACh is released from the nerve terminals, and then activates nicotinic the CA secretion. Based on this fact, the present findings that Rb2 inhibited the CA secretory responses evoked by nicotinic receptor stimulation as well as by membrane depolarization in the adrenal medulla seem to support the fact that, ginsenosides from Panax ginseng can decrease the blood pressure in both experimental animals and hypertensive patients [12,34,35,36]. These experimental results indicate that Rb2-induced inhibitory activity of the CA secretory response evoked by stimulation of nicotinic receptors might contribute at least partly to its hypotensive mechanism. ACh, the physiological presynaptic transmitter at the adrenal medulla, which is released by depolarizing splanchnic nerve terminals and then activates nicotinic receptors, releases the CA, and induces dopamine β-hydroxylase by calciumdependent secretory process [37,38]. In terms of this fact, the present results suggest that Rb2 may inhibit CA secretion evoked by nicotinic stimulation from the splanchnic nerve ending through the blockade of nicotinic receptors. The CA release from the adrenal medulla in response to splanchnic nerve stimulation or nicotinic agonist is mediated by activation of nicotinic receptors located on the chromaffin cells. The exocytotic CA release from the chromaffin cells appears to be essentially similar to that occurring in noradrenergic axons [39,40]. ACh-evoked CA secretion has shown to be caused through stimulation of both nicotinic and muscarinic receptors in guinea-pig adrenal gland [41] as well as in the perfused rat adrenal glands [42]. In support of this idea, it has been found that the ginseng saponins, ginsenoside Rg2, a panaxatriol [6] as well as ginsenoside Rg3, a panaxadiol [8] block the nicotinic ACh receptors or the receptor-operated Na+ channels (but not voltage-sensitive Na+ and Ca2+ channels), inhibit Na+ influx through the channels and consequently reduce both Ca2+ influx and the CA secretion in bovine adrenal chromaffin cells.
In this study, Rb2 inhibited the CA secretory responses evoked by ACh, high K+, and DMPP. It suggests that Rb2 can produce the similar effect as in adrenal medulla of the normotensive rats [4] and also in bovine adrenal chromaffin cells [6,8]. In previous study, ginsenoside-Rg3 inhibited both ACh-induced Ca2+ and Na+ influxes in a concentrationdependent manner similar to that observed with the AChevoked CA secretion [8]. However, it had no or only a slight effect on the CA secretion and Ca2+ influx induced by high K+ concentration or veratridine, an activator of the voltage-sensitive Ca2+ or Na+ channels [5,6]. These results strongly suggested that ginsenoside-Rg3 acts on the nicotinic ACh receptor-operated cation channels but not on the voltage-sensitive Ca2+ or Na+ channels. Furthermore, the ginsenoside-Rg3 inhibition was not overcome by increasing the external ACh and Ca2+ concentrations [8], indicating that the inhibitory effect of ginsenoside-Rg3 is distinct from that of the competitive antagonists of the nicotinic ACh receptors, such as trimethaphan [43,44], and that of blockers of the L-type voltage-sensitive Ca2+ channels, which are competitive with external Ca2+ concentrations, such as diltiazem [45]. In fact, the mode of the ginsenoside-Rg3 antagonism was non-competitive with nicotine [8]. Anyway, these results seem to be quite different from those of the present study that Rb2 significantly inhibited the CA secretory responses evoked by cholinergic nicotinic stimulation as well as an activator of the voltage-sensitive Ca2+ or Na+ channels in the perfused model of rat adrenal medulla. The discrepancy is due to the experimental employment of different accounting methodology and different components between the present and previous studies.
In the present study, Rb2 also time-dependently depressed the CA secretory response evoked by Bay-K-8644, which is known to activate L-type voltage-dependent Ca2+ channels [23,46]. This result indicates that Rb2 may inhibit Ca2+ influx through voltage-dependent Ca2+ channels to the rat adrenomedullary cells. In support of this idea, in cultured bovine adrenal medullary cells, nicotinic (but not muscarinic) receptors mediate the Ca2+-dependent CA secretion [47,48]. It has also been known that the activation of nicotinic receptors stimulates the CA secretion by increasing Ca2+ entry through receptor-linked and/or voltage-dependent Ca2+ channels in both perfused rat adrenal glands [42,49] and isolated bovine adrenal chromaffin cells [50,51,52]. Wada and his coworkers [53] have found that the adrenomedullary chromaffin cells have (i) nicotinic receptor-associated ionic channels, responsible for carbachol-induced Na+ influx, (ii) voltage-dependent Na+ channels, responsible for veratridine-induced Na+ influx and (iii) voltage-dependent Ca2+ channels (VDCC), suggesting that the influx of Na+ caused either by carbachol or by veratridine leads to activate voltage-dependent Ca2+ channels by altering membrane potentials, whereas high K+ directly activates voltage-dependent Ca2+ channels without increasing Na+ influx. In the present study, the finding that the CA secretory responses evoked by Bay-k-8644 as well as by high K+ were depressed in the presence of Rb2 indicates that this inhibitory effect of Rb2 is exerted by the direct inhibition of calcium influx through VDCC into the adrenal chromaffin cells. Furthermore, slight elevation in the extracellular potassium concentration increases both the frequency of spontaneous action potentials and the CA secretion [54], suggesting that the influx of calcium that occurs during action potentials is directly linked to the rate of secretion. These findings that Rb2 inhibited the CA secretion evoked by Bay-K-8644 as well as by high K+ suggest that Rb2 can inhibit directly the VDCC. In the bovine chromaffin cells, stimulation of nicotinic, but not muscarinic ACh receptors is known to cause CA secretion by increasing Ca2+ influx largely through VDCC [55,56]. Therefore, it seems that these inhibitory effects of Rb2 on the CA secretion evoked by ACh, DMPP, Bay-K-8644 and veratridine may be mediated by inhibiting Ca2+ influx through voltage-dependent Ca2+ channels due to activation of nicotinic receptor-associated ionic channels, responsible for carbachol-induced Na+ influx, as well as of voltage-dependent Na+ channels, responsible for veratridine-induced Na+ influx.
The present study has also shown that Rb2 inhibits the CA secretion evoked by cyclopiazonic acid. Cyclopiazonic acid is known to be a highly selective inhibitor of Ca2+-ATPase in skeletal muscle sarcoplasmic reticulum [25,26] and a valuable pharmacological tool for investigating intracellular Ca2+ mobilization and ionic currents regulated by intracellular Ca2+ [57]. Therefore, it is felt that the inhibitory effect of Rb2 on the CA secretion may also be associated with the mobilization of intracellular Ca2+ from the cytoplasmic calcium store. This is consistent with the findings obtained in skinned smooth muscle fibers of the longitudinal layer of the guinea-pig ileum, where Ca2+-uptake was also inhibited by cylopiazonic acid [58]. Suzuki and his coworkers [57] have shown that cyclopiazonic acid easily penetrates into the cytoplasm through the plasma membrane and reduces Ca2+-ATPase activity in sarcoplasmic/endoplasmic reticulum, resulting in an increase in the subsequent Ca2+ release from those storage sites.
In conclusion, as shown in Fig. 9, the results of the present study have strongly suggested that Rb2 inhibits the CA secretion by stimulation of cholinergic nicotinic receptors as well as by direct membrane depolarization in the isolated perfused rat adrenal glands. It seems that this inhibitory effect of Rb2 is exerted by blocking influx of Na+ and Ca2+ through ionic channels into the adrenomedullary chromaffin cells as well as by inhibiting the release of Ca2+ from the cytoplasmic calcium store at least partly via the increased NO production due to the activation of nitric oxide synthase. Based on these results, the ingestion of Rb2 may be helpful to prevent or alleviate the cardiovascular diseases, through inhibition of CA secretion from adrenomedullary cells and consequent reduction of the CA level in the circulation.



 XML Download
XML Download