

 PDF
PDF ePub
ePub Citation
Citation Print
Print


ABBREVIATIONS
NBC
nerve blockers cocktail
TEA
tetraethylammonium
APA
apamin
Glib
glibenclamide
L-NAME
NG-nitro-L-arginine methyl ester
ODQ
1H-(1,2,4)oxadiazolo (4,3-A)quinoxalin-1-one
sGC
soluble guanylate cyclase
NO
nitric Oxide
ECL
enterochromaffin-like
GI
gastrointestinal
GPCR
G-protein-coupled receptor
ATR
atropine
KATP channel
ATP-sensitive K+ channel
KBC
K+ channel blockers cocktail
SNP
sodium nitroprusside
[Ca2+]i
intracellular Ca2+
NSCC
nonselective cation channels
ICC
interstitial cells of Cajal
INTRODUCTION
Histamine was first identified as an autacoid that showed strong vasoactive responses. Histamine is a low-molecular-weight amine released by enterochromaffin-like (ECL) cells and plays multiple roles in the gastrointestinal (GI) tract including secretion of gastric acid [1]. However, it is also synthesized in other cells, such as the gastric mucosa, mast cells, basophils, and parietal cells [2,3]. Inflammation and/or gastric ulcers can recruit hematopoietic cells, such as mast cells, to the tunica muscularis and these cells then release several factors, such as histamine, in the stomach.
Histamine is now known to be an important biochemical paracrine mediator and/or neurotransmitter in diverse physiological and pathophysiological conditions in several tissues. It plays various roles via aminergic G-protein-coupled receptor (GPCR) subtypes H1-H4. Histamine is known to participate in immunoreactivity, the regulation of vascular tone, the secretion of gastric acid, allergic, and inflammatory reactions [2,4,5,6]. Because histamine is stored mainly in mast cells, although it is released actively from ECL cells in the GI tract, it is expected to contribute to the regulation of gastric secretion and motility in the GI tract. Each type of histamine receptor (H1-H4) has been identified in the human GI tract. Additionally, the development of highly selective antagonists has enabled characterizing different responses to histamine [4,5,6]. However, especially regarding the regulation of GI smooth muscle motility by histamine, little is known and the situation is complex because histamine shows dual effects on GI contractility. In fact, canine gastric muscularis mucosa produced contraction at low concentrations of histamine but relaxation at higher concentration via H1 and H2 receptors, respectively [7,8]. Recently, species differences in the colon were also reported: histamine has contractile effects in simian colon smooth muscle but causes relaxation in mouse, through different H receptors [9]. Thus, histaminergic effects on GI motility seem to be species- and organ-specific.
We have recently found that high K+ stimulation produced relaxation via nitric oxide (NO) and soluble guanylate cyclase (sGC) in the longitudinal smooth muscle of the human gastric corpus and fundus [10,11]. Because the gastric corpus and fundus is a specific region for acid secretion and receptive relaxation, such as gastric accommodation after meal ingestion, NO-mediated regulatory mechanisms on GI motility and secretion would seem to be important.
In this study, we sought to determine the relaxing effects of histamine in human gastric circular smooth muscle of the corpus. As histamine plays a key role in the secretion of gastric acid in the gastric corpus, coordinated smooth muscle relaxation during secretion might provide more harmonic and smoother processes. Although histamine showed both contraction and relaxation, the relaxing effect was more prominent at submicromolar to micromolar concentration ranges of histamine via an H2 receptors.
Go to : 
METHODS
Tissue preparation for isometric contraction
The experimental protocol for using human stomach was approved by the Institutional Review Board for Clinical Research of Chungbuk National University. The written informed consent was obtained from all patients who donated gastric tissue. Human gastric tissues from greater curvature were obtained from patients who underwent gastrectomy [10,11,12,13]. Specimens from macroscopically normal tissue of neoplastic area were removed immediately after surgical resection of stomach. In Krebs (KRB) solution, specimens were pinned down on sylgard plate. After removal of mucosa and submucosa, muscle strips (0.5×2 cm, 0.5 cm thickness) were prepared from the corpus and fundus according to circular muscle direction and mounted to organ bath (25 ml and 75 ml) of isometric contractile measuring system. For confirmation pathologist identified smooth muscles of human stomach using HE staining. In vertical chamber, one end of strip was tied tightly to the holder and the other side was linked to force transducer by hook type holder (Harvard, USA). Force transducer was connected to PowerLab-Data Acquisition System, which was linked to IBM compatible computer operated by Charter v5.5 software (ADinstruments, Colorado, USA) for measuring isometric contraction. Each strip was stretched passively to resting tension after 1.5~2 hours equilibration. Then contractile responses of the strip to the high K+ (50 mM, 10 min) was repeated two or three times until the responses were reproducible.
Solution and drugs
KRB solution (CO2/bicarbonate-buffered Tyrode) contained (in mM): NaCl 122, KCl 4.7, MgCl2 1, CaCl2 2, NaHCO3 15, KH2PO4 0.93, and glucose 11 (pH 7.3~7.4, bubbled with 5% CO2/95% O2). Equimolar concentration of Na+ was replaced by K+ to make high K+ (50 mM) solution. The external solution was changed by solutions which had previously been incubated (bubbled with 5% CO2/95% O2, 36℃) in water bath before the application. Pretreatment of various blockers was applied for 12~15 min before the application of histamine. K+ channel blockers cocktail (KBC) was applied before application of stimulators to block each K+ channel responses. KBC contained 4-aminopyridine (4-AP, 2 mM), tetraethylammonium, (TEA, 5 mM); apamin (APA, 300 nM); glibenclamide (Glib, 20 µM). When it is required to rule out nerve mediated response, tetrodotoxin (TTX, 0.4 µM) only and/or nerve blockers cocktail (NBC) was used. NBC contained TTX (0.4 µM), guanethidine (1 µM), and atropine (ATR, 1 µM) [10,11]. All drugs used in this study were purchased from Sigma.
Statistics
Data were expressed as means±standard errors of the mean (means±SEM). The Student's t-test was used to measure the statistical significance. p-value less than 0.05 were regarded as statistically significant.
Go to :
RESULTS
Effect of histamine on spontaneous phasic and tonic contractions of circular smooth muscle in human gastric corpus
Circular smooth muscle from human gastric corpus showed spontaneous contraction of 0.2±0.03 g with a frequency of 2.2±0.19 cycles/min (n=68, respectively; Fig. 1). In human stomach, histamine produced both relaxation and contraction. As shown in Fig. 1A, histamine (10 µM) produced tonic relaxation but relaxant effect was returned to basal level in a few minutes. However, transient tonic relaxation was observed in few tested tissues at micro-molar ranges of histamine. Meanwhile, most tested tissues showed tonic relaxation in circular smooth muscle of human gastric corpus in a concentration-dependent and completely reversible manner (Fig. 1B). Histamine produced tonic relaxation from basal tone to -0.04±0.016 g, -0.06±0.018 g, -0.08±0.021 g, -0.11±0.021 g, -0.12±0.019 g, -0.16±0.024 g, -0.14±0.028 g, respectively (at 0.05, 0.1, 0.2, 1, 2, 5, and 10 µM; n=19, 23, 21, 42, 51, 55, and 43, respectively; p<0.05; right panel of Fig. 1A). Application of higher concentration of histamine such as 20, 50, and 100 µM produced slight tonic relaxation of -0.07±0.029 g, -0.05±0.031 g, and -0.03±0.027 g, respectively (n=4, 3, and 3, respectively; data not shown). In several cases, histamine produced dual relaxant and contractile effects simultaneously in the same tissue as shown in Fig. 1C. Histamine enhanced the amplitude of phasic contraction to 241±55.6%, 223±69.3%, 262±78.2%, 306±115%, 441±159.2% of the control (at 0.5, 1, 2, 5, and 10 µM, respectively; n=3, 4, 4, 4, and 2, respectively).
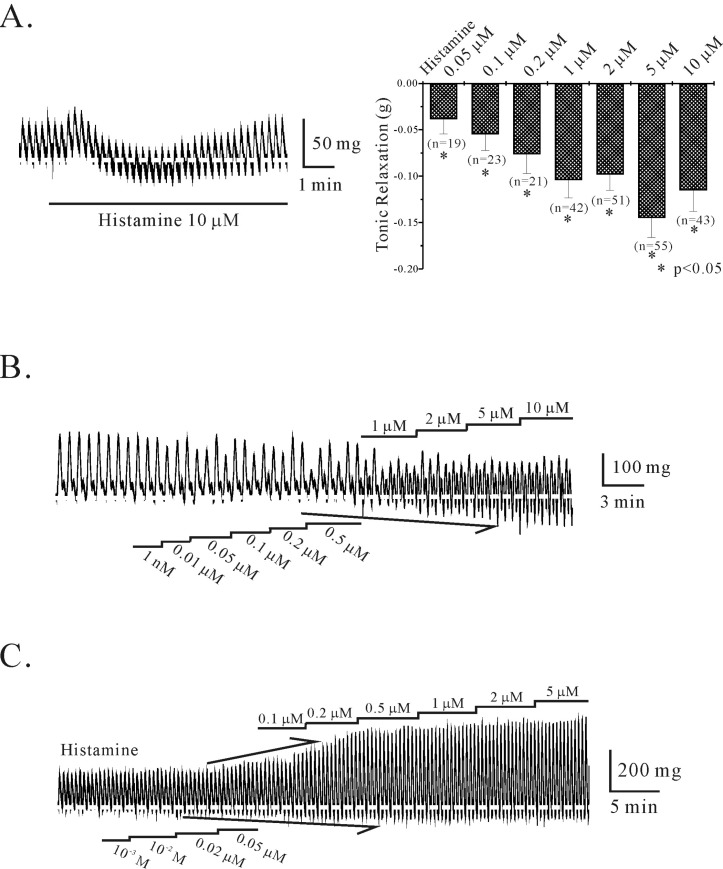 | Fig. 1Effect of histamine on spontaneous phasic and tonic contractions of circular smooth muscle of human gastric corpus. (A) In human gastric corpus, histamine (10 µM) produced tonic relaxation of circular smooth muscle. However, it returned to basal contraction in a few minutes. In right panel, histamine-induced relaxation was summarized. (p<0.05) (B) Most tested tissues showed concentration-dependent tonic relaxation in circular smooth muscle of human gastric corpus. Histamine (1, 2, 5, and 10 µM) produced tonic relaxation from basal tone to -0.11 g, -0.12 g, -0.16 g, and -0.14 g, respectively. (C) In some cases, histamine produced both relaxation and contraction simultaneously in the same tissue. Histamine (0.5, 1, 2, 5, and 10 µM) increased the amplitude of phasic contraction to 241%, 223%, 262%, 306%, and 441% of the control respectively; n=3, 4, 4, 4, and 2, respectively).
|
Effects of tetrodotoxin (TTX) and glibenclamide on histamine-induced tonic relaxation of circular smooth muscle in human gastric corpus
Histamine-induced tonic relaxation was not affected by nerve since pre-treatment of tetrodotoxin (TTX, 0.4 µM) did not affect histamine-induced tonic relaxation (n=3; p>0.05; data not shown). Recently, histamine-induced relaxation was reported to be inhibited by glibenclamide which is known to block ATP-sensitive K+ (KATP) channel in murine colon [9]. Therefore we also studied effect of glibenclamide on histamine-induced tonic relaxation in circular smooth muscle of human gastric corpus. As shown in Fig. 2A and B, histamine-induced tonic relaxation was not affected by pre- and post-application of glibenclamide. In the presence of glibenclamide, histamine produced tonic relaxation by -0.08±0.036 g, -0.1±0.042 g, -0.09±0.043 g, -0.08±0.025 g, -0.08± 0.035 g, -0.09±0.039 g, respectively (at 0.2, 0.5, 1, 2, 5, and 10 µM; n=5, 5, 5, 7, 5, and 5, respectively). Post-application of glibenclamide (20 µM) did not inhibit histamine (10 µM)-induced tonic relaxation (0.2±0.08 g, n=13). These results imply that histamine-induced relaxation was not mediated by nerve and KATP channel in human stomach.
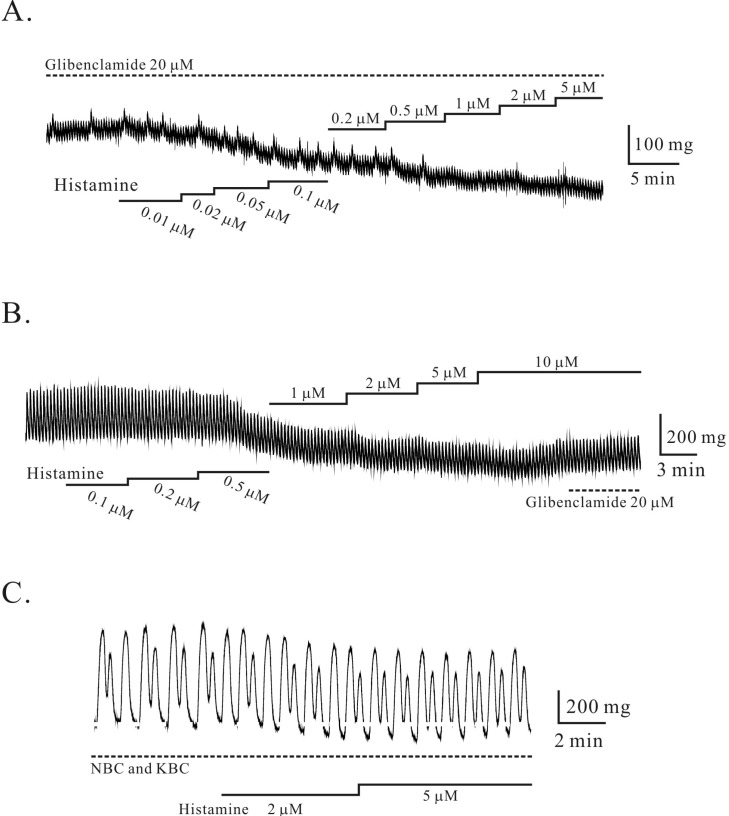 | Fig. 2Histamine-induced relaxation in circular smooth muscle of human gastric corpus. (A, B) Histamine-induced tonic relaxation was not inhibited by pre- and post-application of glibenclamide which block ATP-sensitive K+ (KATP) channel. (C) Pretreatment of nerve blockers cocktail (NBC) and K+ channel blockers cocktail (KBC) did not affected histamine-induced tonic relaxation.
|
Effects of nerve blockers cocktail (NBC), 4-aminopyridine (4-AP), tetraethylammonium (TEA), and apamin (APA) on histamine-induced tonic relaxation in circular smooth muscle of human gastric corpus
To investigate whether K+ channels could be activated by histamine in circular muscle layer, we studied histamine-induced tonic relaxation in the presence of K+ channel blockers cocktail (KBC) and NBC. As shown in Fig. 2C, histamine-induced tonic relaxation was not affected by NBC and KBC (see methods). In the presence of NBC and KBC, histamine produced tonic relaxation of -0.07±0.03 g, -0.13±0.02 g, -0.19±0.03 g, -0.22±0.05 g (at 1, 2, 5 and 10 µM; n=3, 4, 4, 3, respectively). Although data not shown, we also examined whether each K+ channel could be activated by histamine in circular muscle layer. 4-AP (2 mM) produced tonic and phasic contraction of 0.41±0.214 g and 1.55±0.752 g (1.14±0.230 cycles/min; n=13, 13, and 12) [10]. In the presence of 4-AP, histamine produced tonic relaxation by 0.224±0.084 g, 0.37±0.148 g, 0.46±0.207 g, 0.57±0.216 g, respectively (at 1, 2, 5 and 10 µM; n=9, 13, 12, and 10, respectively; data not shown). In the presence of TEA (5 mM), histamine also produced tonic relaxation by 0.09±0.049 g, 0.08±0.021 g, 0.07±0.028 g, respectively (at 1, 2, and 5 µM; n=10, 11, and 10, respectively; data not shown). However, histamine (10 µM) in the presence of TEA produced tonic contraction of 0.05±0.065 g (n=9). Finally, histamine-induced tonic relaxation was studied in the presence of apamin. In the presence of apamin, histamine also produced tonic relaxation by 0.07±0.021g, 0.049±0.018 g, 0.06±0.030 g, 0.077±0.027 g, 0.09±0.033 g, respectively (at 0.5, 1, 2, 5, and 10 µM; n=6, 6, 6, 6, and 4, respectively; data not shown).
Effect of NG-nitro-L-arginine methyl ester (L-NAME) on histamine-induced relaxation of circular smooth muscle in human gastric corpus
The involvement of nitric oxide (NO) and sGC on histamine-induced tonic relaxation of circular smooth muscle in human gastric corpus was investigated. Histamine-induced tonic relaxation was significantly antagonized and/or reversed to contraction by NG-nitro-L-arginine methyl ester (L-NAME), an NO synthesis inhibitor. L-NAME (100 µM) produced tonic and phasic contraction of 0.9±0.12 g and 0.3±0.07 g (n=32 and 23, respectively). In the presence of L-NAME, histamine-induced tonic relaxation was inhibited and/or reversed to tonic contraction by 0.05±0.017 g, 0.08±0.026 g, 0.13±0.034 g, 0.14±0.041 g, respectively (at 1, 2, 5, and 10, respectively; n=31, 22, 22, and 26; Fig. 3A).
 | Fig. 3Effect of NG-nitro-L-arginine methyl ester (L-NAME) on histamine-induced relaxation of circular smooth muscle in human gastric corpus. (A) Histamine-induced tonic relaxation was significantly inhibited by L-NAME. In the presence of L-NAME (100 µM), histamine-induced tonic relaxation was inhibited and/or reversed to tonic contraction. (B) In the presence of ODQ (10 µM) which inhibit soluble guanylyl cyclase (sGC), histamine-induced tonic relaxation was inhibited and/or reversed to tonic contraction.
|
Histamine-induced tonic relaxation was also studied using ODQ which is known to inhibit soluble guanylyl cyclase (sGC). In the presence of ODQ (10 µM), histamine-induced tonic relaxation of human gastric smooth muscle was inhibited and/or reversed to tonic contraction by 0.02±0.008 g, 0.004±0.017 g, 0.05±0.027g, 0.126±0.047 g, respectively (at 0.1, 1, 2, and 5 µM; n=20, 25, 25, and 25, respectively; Fig. 3B).
Effects of ranitidine (H2 receptor blocker) on histamine-induced tonic relaxation of circular smooth muscle in human gastric corpus
To identify specific receptor type of histamine-induced tonic relaxation, the effect of ranitidine was investigated in circular smooth muscle of human gastric corpus. In Fig. 4, ranitidine slightly increased initial tonic contraction. In the presence of ranitidine (0.5 mM), histamine-induced tonic relaxation was significantly suppressed compared to that of control (p<0.05). Histamine produced insignificant effect on motility of circular smooth muscle of human gastric corpus like 0.01±0.017 g, 0.03±0.032 g, 0.02±0.022 g, -0.001±0.031 g, 0.02±0.039 g, 0.02±0.048 g, 0.01±0.055 g, respectively (at 0.1, 0.2, 0.5, 1, 2, 5, and 10 µM; n=11, 6, 11, 11, 11, 11, and 9, respectively).
 | Fig. 4Effect of H2 receptor blockers on histamine-induced relaxation of circular smooth muscle in human gastric corpus. Effects of ranitidine (H2 receptor blocker) on histamine-induced tonic relaxation is shown. In the presence of ranitidine (0.5 mM), histamine-induced tonic relaxation was significantly suppressed compared to that of control (p<0.05).
|
Go to :
DISCUSSION
In this study, we found that histamine-induced tonic relaxation in the circular smooth muscle of the human gastric corpus. Histamine-induced tonic relaxation was completely reversible and blocked by L-NAME (100 µM) and ODQ (10 µM). In addition, H2 blockers inhibited histamine-induced tonic relaxation. These findings suggest that the histamine-induced tonic relaxation of circular smooth muscle from human gastric corpus might be mediated by the activation of H2 receptor and NO/sGC pathways. This is the first report showing histamine-induced tonic relaxation via activation of H2 receptor and NO/sGC pathways in human stomach.
Histamine has diverse physiological and pathophysiological roles, such as in immune/allergic responses, the regulation of vascular tone, and the secretion of gastric acid in the GI tract via its specific receptors, H1-H4 [4,5,6]. During the secretion of gastric acid and inflammation, coordinated gastric motility may be associated with its activity. For example, tonic gastric corporal relaxation by histamine during the secretion of gastric acid will promote coordinated secretion via protection of constriction and/or atropy of gastric folds (Fig. 1A and B, and Fig. 2). In fact, we found high concentration of sodium nitroprusside (SNP; NO-releasing compound, 5 µM) produced inhibition of spontaneous contraction (47% of the control) and basal tone (-1.1 g) in our previous study [12]. However, histamine did not inhibit spontaneous gastric contraction significantly as shown in Fig. 1A and 1B (p>0.05). In this study, we also found histamine produced tonic relaxation (-0.1~-0.16 g) via activation of NO-related pathway but it was significantly different compared to maximal effect by SNP (p>0.05). These results showed that histamine produced tonic relaxation via NO-related pathways but weakly rather than maximum effect by SNP. Since sub-micromolar concentration of SNP inhibits tonic contraction rather than spontaneous gastric contraction in human stomach (unpublished data), it imply that histamine-induced tonic relaxation might be activated via different level of NO-related pathways system compared to SNP-mediated system. Therefore, relaxing patterns and physiological role of SNP and/or histamine in human gastric corporal circular smooth muscle might be different such as histamine which supports acid secretion by tonic relaxation and/or promotes peristaltic movement for secretion from the mucosa to the gastric lumen. Furthermore, we believe that human stomach might have different types of NO-dependent regulating system of gastric motility compared to that of other smooth muscles. Further study must be needed in the future in human gastrointestinal (GI) tract including stomach.
Histamine is well known to regulate blood pressure via potent vasoconstriction in cerebral vascular smooth muscles [14,15,16,17]. However, it also inhibits excitability and/or contractility in other smooth muscles, such as the trachea [18]. Furthermore, a dual action, with contraction and/or relaxation has been reported in the stomach too [7,8,9]. Generally, histamine is known to activate G proteins and modulate cyclic nucleotides [6,15]. Thus, the regulation of cyclic nucleotides and intracellular Ca2+ ([Ca2+]i) might be responsible for excitatory and inhibitory responses of histamine in smooth muscles [6,15]. Additionally, histamine is known to produce depolarization of vascular smooth muscle by activation of nonselective cation channels (NSCC) [19,20]. As shown in Fig. 1C, histamine also enhanced phasic contraction of circular smooth muscle of human gastric corpus. Therefore it might be responsible for activation of these pathways by histamine too. Further study will be needed for elucidation of this mechanism.
Regarding the regulation of GI motility, the effect of histamine on contractility is complex [7,8,9]. There are species- and concentration-dependent differences in histamine-induced effects in several organs [18]. In this study, we also found histamine-induced contraction and relaxation. Sometimes both effects were observed even in the same tissue of the human stomach (Fig. 1C). In canine stomach, histamine showed dual effects on the contractility of the gastric muscularis mucosa, such as contraction and relaxation via H1 and H2 receptors, by high (>10 µM) and low (<10 µM) concentrations, respectively [7]. Furthermore, three types of responses by histamine on contractility-contraction, contraction followed by relaxation, and relaxation-were reported in ruminant stomach [8]. Recently, a contractile effect of histamine was reported in simian colonic smooth muscle, but relaxation was observed in mouse, involving different types of H receptors. Generally, excitatory and inhibitory effects of histamine on gastric motility, and on other smooth muscles, have been identified using antagonists of H1 and H2 receptors including H3 receptor-mediated relaxation, respectively [7,8,9,18]. However, we found that it is also hard to dissociate the contractile vs relaxant responses to specific concentrations of histamine. Therefore, this study was focused to relaxant effect by histamine first in human stomach.
In the GI tract, including humans, H1 and H2 play major physiological functions rather than H3 and H4 [3,7,8]. Histamine H1 receptor have been reported to be expressed in myenteric nerve plexus of the human intestine [21]. In contrast, H2 receptors appear to be expressed on parietal cells in the fundus [22] and they have also been found on immune cells and the myenteric plexus in humans [21]. However, the role of H3 and H4 is not well studied and there are some controversies compared with that of the H1 and H2 receptors [23,24].
Recently, species differences in histamine-induced responses were reported in murine and simian colons. Histamine produced contraction via activation of H1 and H4 receptors in the simian colon but it produced relaxation via activation of H2 receptors in mouse [9]. In Fig. 4, pretreatment with ranitidine (0.5 mM) significantly inhibited histamine-induced tonic relaxation in the circular smooth muscle of the human gastric corpus. Because H2 receptor antagonists, such as ranitidine, are known to inhibit histamine-induced relaxation of canine and ruminant stomach, it is possible that H2 receptors may also be involved in histamine-induced relaxation in the human gastric corpus [7,8]. Furthermore, H2 receptor may be one of the targets for clinical application in functional dyspepsia because we found that ranitidine inhibited histamine-induced tonic relaxation in the human stomach (Fig. 4). H2 receptor antagonists such as ranitidine may possibly have a therapeutic effect of around 20% over placebo. Functional dyspepsia (and/or non-ulcer dyspepsia) is a diagnosis of exclusion based on dyspeptic symptoms in the absence of structural abnormalities on endoscopic observation [25]. Since H2 receptor antagonists mainly improve functional dyspepsia in upper gastric pain, antagonistic effect of ranitidine on tonic relaxation of corporal human stomach and its associated vessel contractility might reform stomach to more comfortable state.
Generally, activation of K+ channels opposes membrane excitability then inhibits smooth muscle contractility [10,17,25,26]. To rule activation of K+ channel during the application of histamine, the effects of pretreatment with KBC before the application of histamine were also studied. In particular, it has been reported the activation of KATP channel by histamine was responsible for histamine-induced relation in murine colon [9]. However, as shown in Fig. 2A and 2B, pre- and post-application of glibenclamide did not inhibit histamine-induced relaxation as well as KBC (Fig. 2C).
In the human stomach, nNOS neurons are present and NO is localized in the myenteric plexus [27]. Additionally, NO partly mediates relaxation, by graded distention in the human gastric fundus [28]. As shown in Fig. 3A and 3B, histamine-induced tonic relaxation was inhibited and/or converted to contraction by pretreatment with L-NAME (100 µM) or ODQ (10 µM) except in two cases, respectively. These findings indicate that NO-mediated pathways may be involved in histamine-induced tonic relaxation in the human stomach. Since histamine also produced tonic relaxation via activation of NO pathway, histamine might play an important role for adaptive relaxation in human stomach too [10,11,12]. As shown in the Results, histamine produced both contraction and relaxation in the same tissue, as shown in the circular smooth muscle of corpus (Fig. 1C). In the presence of L-NAME and ODQ, however, histamine-induced tonic relaxation was antagonized and/or reversed to contraction in both tissues (Fig. 3A and 3B). In our previous study, in fact, we reported activation of the NO/sGC pathways in high K+-induced relaxation in longitudinal smooth muscle of the human gastric corpus and fundus [10,11]. It has also been reported that interstitial cells of Cajal (ICC) express NO-sensitive sGC and it is the primary target of NO released from nNOS neurons even in human stomach [13,29,30,31,32,33]. Because NO signals are transmitted to smooth muscle of the GI tract and ICC in human stomach was already reported, a NO-mediated pathway may be responsible for histamine-induced relaxation in the human stomach [13,32]. The underlying cellular mechanisms of action of histamine and distribution of the related H receptors in the human stomach need be studied further.
Go to :
 XML Download
XML Download