

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Histamine exerts a variety of physiological functions and the activation of histamine receptors by several histamine receptor agonists produces a several pharmacological effects [1,2]. A several lines of evidence suggest that one of the important functions of histamine system is the regulation of the blood glucose level. For example, treatment with a combination histamine H1 and H2 receptor antagonist lowers the blood glucose level in diabetic patients [3]. In addition, Schusdziarra et al have demonstrated that histamine H2 receptor participates in gastrogenic hyperglycemia and hyperinsulinemia in dogs [4]. Moreover, Lal has previously reported that histamine H1 receptor antagonists administered orally produce differential actions in the regulation of the blood glucose level in a clinical study [5]. For example, certrizine administered orally causes a slight elevation of the blood glucose level, whereas clemastine, terfenadine or astemizole administered orally causes a fall of the blood glucose level during fasting. The pretreatment with antihistamine administered systemically blocks the hypoglycemic effect induced by endotoxin shock [6]. However, an earlier study has reported that plasma glucose levels are not affected by histamine or cimetidine [7].
In addition to the direct involvement of histamine receptors in the regulation of blood glucose level, several studies have demonstrated that activation or inactivation of histamine receptors appear to exert a modulatory role in the regulation of the blood glucose level. For example, pretreatment with the histamine H1 receptor antagonist (diphenhydramine and promethazine) or histamine H2 receptor antagonist (cimetidine) partially blocks systemically administered neurotensin-induced hyperglycemia [8]. In addition, histamine H1 receptor antagonist (pyrilamine) administered into the third ventricle suppresses hyperglycemia induced by intraventricular injection of neostigmine, whereas histamine H2 receptor antagonist (ranitidine) administration into the third ventricle enhances neostigmine-induced hyperglycemia [9]. Moreover, oral administration of histamine H2 receptor antagonists such as cimetidine or ranitidine potentiates the hypoglycemic response to glipizide in diabetic patients [10]. H2 receptor antagonist enhances alcohol-induced hypoglycemia [11]. However, intravenous administration of cimetidine does not affect the insulinogetic effect of glucose [12]. Furthermore, systemic treatment with H2 receptor antagonist (metiamide) or intracerebroventricular (i.c.v.) pretreatment with H2 receptor antagonist (cimetidine) shows no modulatory effect on clonidine-induced hyperglycemia [13,14].
In addition to the involvement of histamine systems in the peripheral system, several lines of evidence have suggested that histamine system located in the brain also appear to be involved in the regulation of blood glucose level. For example, Nishibori et al have demonstrated that brain is an important site for the regulation of blood glucose level [15]. They have demonstrated that i.c.v. administration of histamine increases the plasma glucose level.
Although the involvement of histamine receptors located at the peripheral and brain sites in the regulation of the blood glucose level has been well demonstrated in numerous previous studies, the possible roles of histamine receptors located in the spinal cord for the regulation of the blood glucose level have not been well characterized yet. Thus, in the present study, the effects of various types of histamine receptor agonists or antagonists administered spinally on the blood glucose level were examined in control and D-glucose-fed mice.
Go to : 
METHODS
These experiments were approved by the Hallym University Animal Care and Use Committee (Registration Number: Hallym 2009-05-01). All procedures were conducted in accordance with the 'Guide for Care and Use of Laboratory Animals' published by the National Institutes of Health.
Experimental animals
Male Hsd: CD-1 (ICR) [Charles River, USA] mice, weighing 24~26 g, were used for all the experiments. Five mice were housed per cage in a room maintained at 22±0.5℃ with an alternating 12 h light-dark cycle. Food and water were available ad libitum. The animals were allowed to adapt to the laboratory for at least 2 h before testing and were only used once. Experiments were performed during the light phase of the cycle (10:00~17:00). The animals were fasted for 16 hours.
Oral and intrathecal (i.t.) injection
Oral administration was performed with gavage in a volume of 1 ml/kg body weight. I.t. administration was performed in conscious mice, following the method of Hylden and Wilcox, using a 30-gauge stainless-steel needle attached to a 25 µl Hamilton microsyringe [16]. The mice were gently handled and i.t. injection was performed as quickly as possible so that the mice can avoid the stress as much as possible. The i.t. injection volume was 5 µl and the injection site was verified by injecting a similar volume of 1% methylene blue solution and determining the distribution of the injected dye in the spinal cord. The dye injected i.t. was distributed both rostrally and caudally but with short distance (about 0.5 cm) and no dye was found in the brain. The success rate for the injections was consistently found to be over 95%, before the experiments were done.
Drugs
2-pyridylethylamine, cetirizine, dimaprit, ranitidine, α-methylhistamine, carcinine, VUF 8430 and JNJ 7777120 were purchased from Tocris Bioscience Co. (Minneapolis, MN, USA). D-glucose was purchased from Sigma Chemical Co. (St. Louis, MO, USA). 2-pyridylethylamine, cetirizine, dimaprit, ranitidine, α-methylhistamine, carcinine and VUF 8430 were dissolved in saline. JNJ 7777120 was prepared following steps: (A) 1 g of JNJ 7777120 was dissolved in 0.5 ml of ethanol plus 0.5 ml of polyethylene glycol 400. (B) Separately, 100 mg of sodium carboxymethylcellulose was dissolved in 9 ml of distilled water. (C) Finally, Solution (A) and Solution (B) were vigorously mixed. This solution (PEC) excluding JNJ 7777120 was used as vehicle control. All drugs were prepared just before use. Blood glucose meter, lancing device and strips were purchased from Roche Diagnostics (Sandhofer Strasse, Mannheim, Germany).
Measurement of blood glucose level
Blood glucose measurements were obtained using blood samples collected by lateral tail vein laceration. A minimum volume (1 µl) of blood was collected as quickly as possible. Glucose level was measured using Accu-Chek Performa blood glucose monitoring system (Sandhofer Strasse, Mannheim, Germany).
Treatment of drugs
At first, mice were treated i.t. with either saline, PEC and histamine receptors (2-pyridylethylamine (0.1~1.0 µg/5 µl), cetirizine (0.05~0.5 µg/5 µl), dimaprit (1~10 µg/5 µl), ranitidine (5~20 µg/5 µl), α-methylhistamine (0.5~5.0 µg/5 µl), carcinine (1~10 µg/5 µl), VUF 8430 (0.5~5.0 µg/5 µl) and JNJ 7777120 (1~10 µg/5 µl)) 10 min before i.t. administration of vehicle as a control or a fixed dose of agonists (phenylephrine, clonidine, dubutamine and terbutaline).
Statistical analysis
The statistical significance of differences between groups was assessed with one-way ANOVA with Bonferroni's post-hoc test using GraphPad Prism Version 4.0 for Windows XP (GraphPad Software, San Diego, CA, USA). The p-values less than 0.05 were considered to indicate statistical significance. All values were expressed as the mean±S.E.M. In our study, we established the mean blood glucose value of the control group through many experiments under matching conditions. Selected mice of established blood glucose level were then used in replication experiments.
Go to :
RESULTS
Effect of hitamine receptor agonists and antagonists on the blood glucose level
Mice were intrathecally (i.t.) administered with 2-pyridylethylamine (from 0.1 to 1 µg), cetirizine (from 0.05 to 0.5 µg), dimaprit (from 1 to 10 µg), ranitidine (from 5 to 20 µg), α-methylhistamine (0.5 to 5 µg), carcinine (from 1 to 10 µg), VUF 8430 (from 0.5 to 5 µg), or JNJ 7777120 (from 1 to 10 µg), and the blood glucose level was measured at 30, 60 and 120 min after i.t. administration. As shown in Fig. 1A (F=0.09607; p=0.9607), 1B (F=0.2140; p=0.8848), 2A (F=0.1932; p=0.8990), 2B (F=0.04422; p=0.9870), 4A (F=0.3528; p=0.7880) and 4B (F=0.6188; p=0.6161), 2-pyridylethylamine, cetirizine, dimaprit, ranitidine, VUF 8430, or JNJ 7777120 did not affect the blood glucose level. I.t. injection with α-methylhistamine, but not carcinine, slightly caused an elevation of the blood glucose level (Fig. 3A (F=0.04059; p=0.9885) and 3B (F=0.2200; p=0.8807)).
 | Fig. 1Effect of 2-pyridylethylamine or cetirizine administered i.t. on the blood glucose level. Mice were intrathecally (i.t.) administered with 2-pyridylethylamine (from 0.1 to 1 µg) (A) or cetirizine (from 0.05 to 0.5 µg) (B). The 16 h fasted mice were pretreated i.t. with 2-pyridylethylamine (C) or cetirizine (D) for 10 min and then, D-glucose (2 g/kg body weight) administered orally once. The blood glucose level was measured at 30, 60 and 120 min after i.t. or oral administration. The blood was collected from tail-vein. The vertical bars indicate the standard error of mean. Each quantified result was analyzed by one-way ANOVA with a Bonferroni post hoc test (C and D: **p<0.01, ***p<0.005; compared to D-Glucose+Saline group). The number of animals used for each group was 8~10.
|
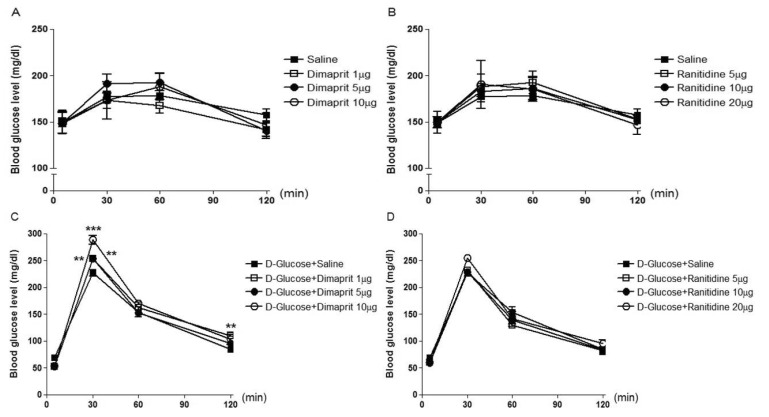 | Fig. 2Effect of dimaprit or ranitidine administered i.t. on the blood glucose level. Mice were intrathecally (i.t.) administered with dimaprit (from 1 to 10 µg) (A) or ranitidine (from 5 to 20 µg) (B). The 16 h fasted mice were pretreated i.t. with dimaprit (C) or ranitidine (D) for 10 min and then, D-glucose (2 g/kg body weight) administered orally once. The blood glucose level was measured at 30, 60 and 120 min after i.t. or oral administration. The blood was collected from tail-vein. The vertical bars indicate the standard error of mean. Each quantified result was analyzed by one-way ANOVA with a Bonferroni post hoc test (C: **p<0.01, ***p<0.005; compared to D-Glucose+Saline group). The number of animals used for each group was 8~10.
|
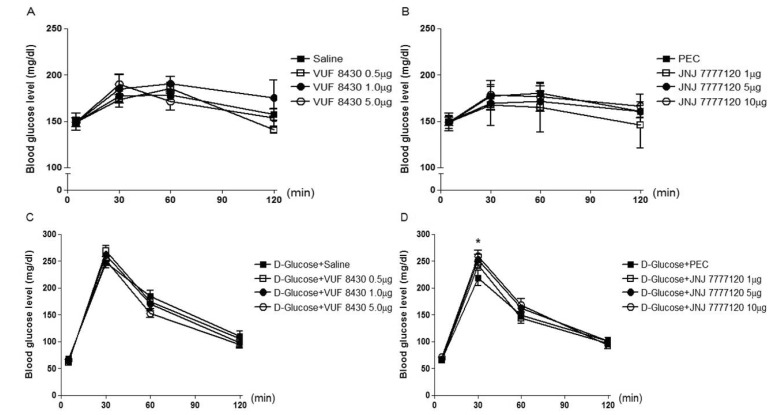 | Fig. 4Effect of VUF 8430 or JNJ 7777120 administered i.t. on the blood glucose level. Mice were intrathecally (i.t.) administered with VUF 8430 (from 0.5 to 5 µg) (A) or JNJ 7777120 (from 1 to 10 µg) (B). The 16 h fasted mice were pretreated i.t. with VUF 8430 (C) or JNJ 7777120 (D) for 10 min and then, D-glucose (2 g/kg body weight) administered orally once. The blood glucose level was measured at 30, 60 and 120 min after i.t. or oral administration. The blood was collected from tail-vein. The vertical bars indicate the standard error of mean. Each quantified result was analyzed by one-way ANOVA with a Bonferroni post hoc test (D: *p<0.05; compared to D-Glucose +PEC group). The number of animals used for each group was 8~10.
|
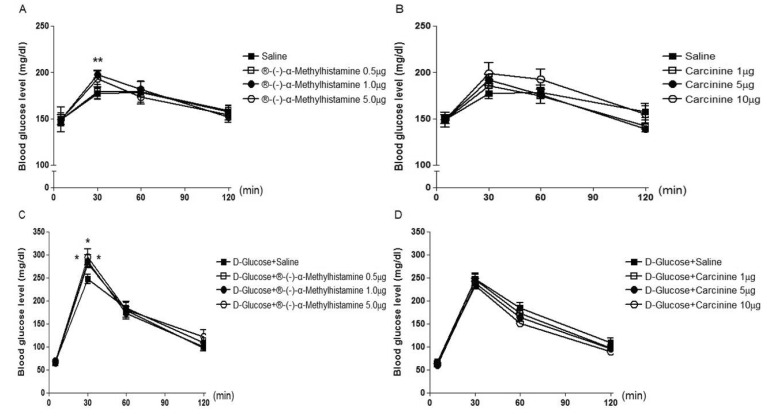 | Fig. 3Effect of α-methylhistamine or carcinine administered i.t. on the blood glucose level. Mice were intrathecally (i.t.) administered with α-methylhistamine (0.5 to 5 µg) (A) or carcinine (from 1 to 10 µg) (B). The 16 h fasted mice were pretreated i.t. with α-methylhistamine (C) or carcinine (D) for 10 min and then, D-glucose (2 g/kg body weight) administered orally once. The blood glucose level was measured at 30, 60 and 120 min after i.t. or oral administration. The blood was collected from tail-vein. The vertical bars indicate the standard error of mean. Each quantified result was analyzed by one-way ANOVA with a Bonferroni post hoc test (A: **p<0.01; compared to saline group, C and D: *p<0.05; compared to D-Glucose+Saline group). The number of animals used for each group was 8~10.
|
Effect of histamine receptor agonists and antagonists on the blood glucose level in D-glucose-fed model
Mice were intrathecally (i.t.) pretreated with 2-pyridylethylamine (from 0.1 to 1 µg), cetirizine (from 0.05 to 0.5 µg), dimaprit (from 1 to 10 µg), ranitidine (from 5 to 20 µg), α-methylhistamine (0.5 to 5 µg), carcinine (from 1 to 10 µg), VUF 8430 (from 0.5 to 5 µg), or JNJ 7777120 (from 1 to 10 µg) for 10 min and D-glucose (2 g/kg) was orally fed, and the blood glucose level was measured at 30, 60 and 120 min after D-glucose administration. As shown in Fig. 1, 2, 3, and 4, in D-glucose-fed model, i.t. pretreatment with cetirizine enhanced the blood glucose level, whereas 2-pyridylethylamine did not affect the blood glucose level (Fig. 1C (F=0.01885; p=0.9963) and 1D (F=0.1807; p=0.9074)). The i.t. pretreatment with dimaprit, but not ranitidine, enhanced the blood glucose level in D-glucose-fed model (Fig. 2C (F=0.04018; p=0.9887) and 2D (F=0.02038; p=0.9958)). In addition, α-methylhistamine, but not carcinine, slightly but significantly enhanced the blood glucose level in D-glucose-fed model (Fig. 3C (F=0.01836; p=0.9964) and 3D (F=0.03596; p=0.9904)). Finally, i.t. pretreatment with JNJ 7777120, but not VUF 8430, slightly but significantly enhanced the blood glucose level in D-glucose-fed model (Fig. 4C (F=0.01462; p=0.9974) and 4D (F=0.03103; p=0.9923)).
Go to :
DISCUSSION
Several earlier studies have demonstrated that the blood glucose level is regulated by histamine receptor agonists or antagonists administered supraspinally or systemically [15,17]. Various studies have demonstrated that histamine H1 and H3 receptors are expressed in the gray matter, especially in the superficial layer and the lamina II area [18,19], whereas histamine H2 and H4 receptors are expressed in the motor neurons in the ventral horn of the spinal cord [20,21]. However, the role of the spinally located histamine receptors in the regulation of the blood glucose level has not been characterized. Thus, the effects of several types of histamine receptor agonists and antagonists administered spinally on the blood glucose level were examined in the present study. We found for the first time that i.t. treatment with α-methylhistamine (a histamine H3 receptor agonist), slightly caused an elevation of the blood glucose level. However, we found that 2-pyridylethylamine (a histamine H1 receptor agonist), cetirizine (a H1 receptor antagonist), dimaprit (a histamine H2 receptor agonist), ranitidine (a histamine H2 receptor antagonist), carcinine (a histamine H3 receptor antagonist), VUF 8430 (a histamine H4 receptor agonist) or JNJ 7777120 (a histamine H4 receptor antagonist) did not affect the blood glucose level. In an earlier study, supraspinal administration with histamine H3 receptor agonist (proxyfan) significantly reduces glucose level compared with vehicle-treated mice [22]. These findings suggest that activation of spinally located histamine H3 receptors appear to play an important role in the regulation of the blood glucose level. In contrast to the results with involvement of histamine H3 receptors in the regulation of the blood glucose level, histamine H1, H2, and H4 receptors located in the spinal cord appear not to be involved in the regulation of the blood glucose level. These results are not in line with previous findings that the selective H3 receptor agonist, immepip, administered to diabetic rats does not alter plasma glucose level [23].
We further examined the possible modulatory roles of spinally located histamine receptors for the regulation of the blood glucose level in D-glucose-fed mice model. In an attempt to investigate this possibility, we examined effects of histamine receptor agonists or antagonists pretreated spinally on the blood glucose level in D-glucose-fed mice. We found in the present study that cetirizine pretreated spinally enhances the blood glucose level, whereas 2-pyridylethylamine does not affect the blood glucose level in D-glucose-fed mice, suggesting that the blockade of histamine H1 receptors located at the spinal cord exerts a modulatory effect on the blood glucose regulation in a potentiative manner in D-glucose-fed model. In addition, we found in the present study that spinal pretreatment with dimaprit, but not ranitidine, enhances the blood glucose level in D-glucose-fed model, suggesting that activation of spinal histamine H2 receptors exert a modulatory effect on the blood glucose regulation in a potentiative manner in D-glucose-fed model. Furthermore, we found in the present study that α-methylhistamine, but not carcinine, pretreated spinally enhances the blood glucose level in an additive manner in D-glucose-fed model, suggesting that activation of spinal histamine H2 receptors does not exerts a modulatory effect on the blood glucose regulation in D-glucose-fed model. Finally, spinal pretreatment with VUF 8430 slightly but significantly reduces the blood glucose level, whereas JNJ 7777120 pretreated spinally slightly but significantly enhances the blood glucose level in D-glucose model, suggesting that activation or the blockade of spinal histamine H4 receptors exert a modulatory effect on the blood glucose regulation in D-glucose-fed model. Taken together, various types of histamine receptors located at the spinal cord are actively exert modulatory roles in the regulation of the blood glucose level in D-glucose-fed model. Inactivation of histamine H1 and H4 receptors located in the spinal cord plays an important role for the modulatory down-regulation of the blood glucose level in D-glucose fed model, whereas activation of spinal histamine H2 receptors may be responsible for modulatory up-regulation of the blood glucose level in D-glucose-fed model. However, histamine H3 receptor does not appear to exert a modulatory role for the regulation of the blood glucose level in D-glucose-fed model. The exact modulatory mechanisms involved in up-regulation or down-regulation of the blood glucose level by histamine receptor receptors activation or inactivation in the spinal cord remains to be clarified further in the future study.
The present study supports the hypothesis that the spinal cord plays an important site for the regulation of the blood glucose level. In an earlier clinical study, Sala et al. have reported that insulin is used to regulate the up-regulation of the blood glucose level in patients with spinal cord injury [24]. In addition, we found recently that substance P or several pro-inflammatory cytokines such as TNF-α, IFN-γ and IL-1β administered spinally causes an elevation of the blood glucose level [25,26]. Taken together from the results of our present study and other groups, more pharmacological studies on the role of the spinal cord for the regulation of the blood glucose level should be further performed in the future.
Go to :
 XML Download
XML Download