

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Excitation of the peripheral vestibular receptors by postural changes induces functional changes to many components of the cardiovascular system, including blood pressure, pulse rate, baroreceptor reflex, and blood flow to the extremities [1]. In addition, cardiovascular function is affected by stimulation of the vestibular nerve and vestibular nuclei [2,3]. Electrical or selective natural stimulation of the vestibular receptors elicits an increase in sympathetic nerve activity [4,5,6]. A postural change from a lying position to a standing position induces a downward fluid shift by increasing the hydrostatic pressure gradient, which produces reduced venous return and cardiac output, consequently reducing arterial pressure. This reduction in arterial pressure evokes a baroreflex response, and arterial pressure stabilizes [7]. In addition, postural changes stimulate the vestibular receptors, which are also thought to be involved in regulating arterial pressure [8], but loss of vestibular function impairs the ability to compensate for orthostatic hypotension [9].
Previous anatomic and physiologic studies indicated interactions between the vestibular nuclei and sympathetic centers in the brainstem. Direct projections from the inferior vestibular nuclei and caudal medial vestibular nuclei to both the nucleus tractus solitaries (NTS) and the dorsal motor nucleus of the vagus nerve [10,11] suggest that vestibular inputs converge with visceral sensory inputs in the NTS, and indicate vestibular influences on the brainstem autonomic response, including cardiovascular control during head movements and motion sickness [12].
Changes in arterial blood pressure resulting from hemorrhage, cardiovascular disorders, or postural changes elicit an autonomic reflex response through baroreceptors. Afferent signals from the baroreceptors are transmitted by the carotid sinus nerve and aortic depressor nerve to the NTS, which is the primary termination region of baroreceptor afferents and the center of interpretation of the context of input signals [13,14]. Increased arterial blood pressure evokes the discharge of baroreceptors in the carotid sinus and aortic arch, and these signals are then integrated with the vestibular signals in the NTS [15]. However, the quantitative roles of the baroreceptor and vestibular receptor in the regulation of blood pressure are poorly understood.
Extracellular regulated protein kinase (ERK) is involved in a complex intracellular signaling cascade that controls various neurobiological effects. ERK is activated via phosphorylation in neurons in response to various excitatory stimuli such as membrane depolarization, glutamate, and electrical stimulation. The active form of ERK, phosphorylated ERK protein (pERK), is crucial for controlling the transcription of immediate early genes such as c-fos [16]. We used immunohistochemical labeling of pERK as a means of identifying the populations of neurons within the vestibular nuclei following deafferentation of the vestibular system [17]. Moreover, previous studies in our laboratory showed that sodium nitroprusside (SNP)-induced acute hypotension increases expression of c-Fos protein [18] and pERK [19] in the vestibular nuclei, and that these effects are abolished by removal of peripheral vestibular receptors.
Our current immunohistochemical studies in the vestibular nuclei are based largely on experiments in anesthetized animals. Since anesthesia can greatly affect the responses of central neurons to physiological stimuli, such studies provide only a limited understanding of the central mechanisms for the vestibular function in normal conscious animals. In this study, to clarify the role of vestibular receptors and baroreceptors on the regulation of blood pressure during acute hypotension, the expression of pERK in the NTS was analyzed in conscious rats that received bilateral labyrinthectomy (BL) and/or baroreceptor unloading via sinoaortic denervation (SAD).
Go to : 
METHODS
Animals
Adult male Sprague-Dawley rats (Changchun, China) weighing 220~250 g were used in this study. The animals had free access to food and water. Efforts were made to minimize the number of animals used and suffering. Experimental animals were divided into four groups for immunohistochemical analysis (n=8/group): a sham group, in which both the sinoaortic baroreceptors and vestibular end organs were intact; the BL group, in which the sinoaortic baroreceptors were intact, but a bilateral labyrinthectomy was performed; the SAD group, in which the sinoaortic baroreceptors were denervated, but the vestibular end organs remained intact; and the SAD+BL group, in which both the sinoaortic baroreceptors and bilateral vestibular end organs were removed. The Institutional Ethical Committee of Wonkwang University on the Experimental Use of Animals approved all the procedures used.
Labyrinthectomy
A chemical labyrinthectomy was performed as described previously [20]. Briefly, after anesthesia with isoflurane (Ilsung Co., Seoul, Korea), 100 µL of sodium arsanilate (100 mg/mL) was intratympanically injected into the bilateral middle ear of the rats, which chemically destroyed the membranous labyrinth. The destruction of epithelial cells in peripheral vestibular receptors was confirmed by confocal microscope observations after rhodamine-palloidin staining (Fig. 1). For rhodamine-palloidin staining, the tissue was sectioned to a thickness of 30 µm on a freezing microtome (Leica; Germany), fixed with 1~2% paraformaldehyde in 0.1 M phosphate-buffered solution (PBS) at room temperature for 15 min, washed with 0.1 M glycine in PBS for 1 min, incubated with 0.1% Triton X-100 in PBS for 1 min, and then incubated with rhodamine-palloidin solution (diluted 1:100; Molecular Probes; USA) in PBS for 15 min. After incubation, the tissue was rinsed 3 times for 5 min each time with PBS. The tissue was mounted for confocal microscopy using PBS-buffered 50% glycerol as mounting medium.
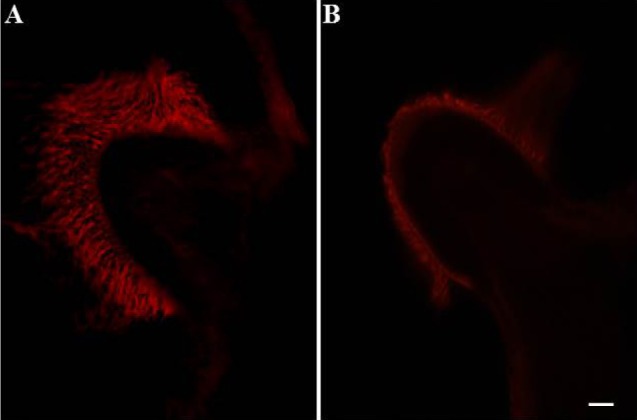 | Fig. 1Confocal microscopic findings in rhodamine-pallodin stained samples showing the effect of arsanilate on vestibular hair cells. Cilia in hair cells disappeared after treatment with arsanilate. (A) Ampulla of the posterior semicircular canal in control; (B) ampulla of the posterior semicircular canal 48 hours after treatment with arsanilate. Magnification=×400; scale bar=10 µm.
|
As a control, saline, instead of sodium arsanilate solution, was injected intratympanically in the sham and SAD rats. The labyrinthectomies were performed 48 hours prior to experimentation.
Sinoaortic denervation
Carotid and aortic sinus denervation was performed as described previously [21]. After anesthesia with isoflurane, the carotid sinus nerve was sectioned bilaterally following a midventral incision in the neck, and the internal, external, and common carotid arteries were stripped of connective tissue at the level of bifurcation and painted with 10% phenolethanol to denervate the carotid sinus. For aortic arch denervation, the aortic arch nerve was severed bilaterally proximal to its junction with the vagus nerve. In the sham and BL groups, rats received similar cervical incisions leaving nerves, vessels, and baroreceptors intact. After the surgery, animals were breathing spontaneously without significant changes in respiratory rhythm. SAD was performed 24 hours prior to experimentation.
Acute hypotension
Two heparinized polyethylene tubes were inserted into the femoral artery for recording the blood pressure, and into the femoral vein for SNP infusion, under isoflurane anesthesia. The tubes were guided toward the skull percutaneously, fixed into the skull, and connected to the tubes of a cybernation metabolism cage to allow free movement in a conscious state during the experiment. The blood pressure was recorded from the unilateral femoral artery using a pressure transducer and physiography (Grass model 7400; USA). SNP was infused in 3 min at a dose of 15 µg/kg/min, and blood pressure decreased by 30~40 mmHg during this period.
Immunohistochemistry
After deep anesthesia with an intraperitoneal injection of sodium pentobarbital (100 mg/kg), animals were sacrificed for immunohistochemical analysis of ERK at 5 (n=8), 10 (n=8), 20 (n=8), or 40 min (n=8) following administration of SNP. We selected the aforementioned time points on the basis of results of earlier studies, in which the kinetics of ERK activation in the brain or spinal cord were reported to be very rapid [16]. The rats were perfused, fixed with 4% paraformaldehyde, and the sucrose-embedded brain stem was sectioned on a cryostat. After nonspecific binding sites were blocked with normal goat serum, primary anti-rabbit polyclonal anti-ERK 1/2 antibody (1:1000 dilution) (Cell Signaling Technology; MA, USA) was applied to tissue sections overnight at 4℃. Thereafter, tissue sections were incubated with a secondary antibody and then with an avidin-biotin complex. The bound complex was visualized by incubating the tissue with diaminobenzadine plus H2O2. Sections were then dehydrated, cleared in xylene, and cover-slipped with Permount (Fisher Scientific; Pittsburgh, PA, USA). For quantification, pERK 1/2-immunopositive neurons in the NTS were counted using a digital image analysis system (Image-Pro; Media Cybernetics; MD, USA) at three different regions in a rostral-to-caudal continuum [18,22].
Data analysis
All data are expressed as mean±standard error. Differences were identified using multivariate analysis of variance. p<0.05 was considered statistically significant.
Go to :
RESULTS
Effect of acute hypotension on expression of pERK-immunoreactive neurons
In control rats with intact labyrinths and baroreceptors, a few pERK-immunoreactive neurons appeared in the NTS following saline infusion (2.0±0.4). However, SNP-induced acute hypotension significantly increased the expression of pERK-immunoreactive neurons in the NTS. Expression of pERK-immunoreactive neurons was observed at the rostral-caudal regions of the NTS, but was mainly increased in the caudal portion of the NTS. The number of pERK-immunoreactive neurons was 125.6±6.4, 243.3±7.5, 21.3±1.4, and 3.3±0.4 at 5, 10, 20, and 40 min after administration of SNP, respectively. The expression peaked at 10 min after administration of SNP and then decreased gradually thereafter (Figs. 2 and 3).
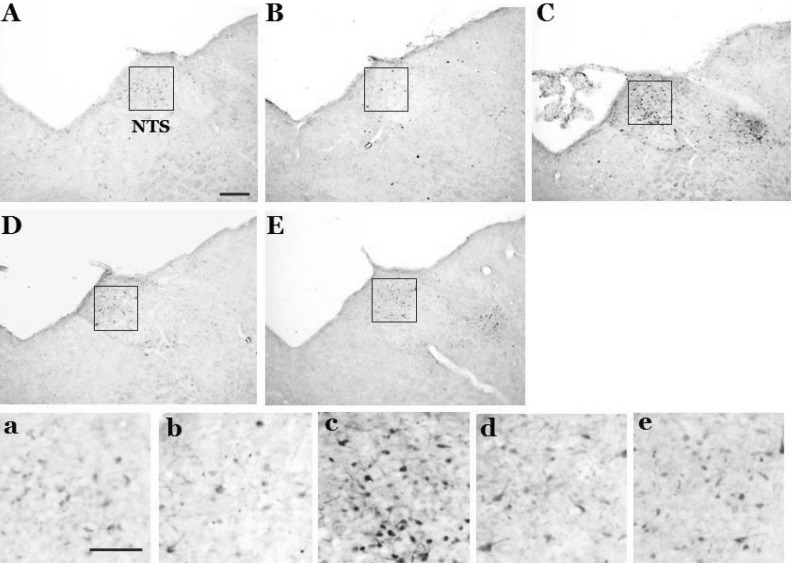 | Fig. 2Photomicrographs showing the effect of acute hypotension on pERK expression in the nucleus tractus solitarius following a 30% reduction in blood pressure that was induced by SNP infusion in conscious rats with intact labyrinths and baroreceptors. Expression of pERK was measured 5 (B), 10 (C), 20 (D), and 40 (E) min after SNP infusion. Control (A) received intravenous administration of saline at the same volume as SNP; NTS, nucleus tractus solitarius (the coordinate: 0.2 mm rostral to the obex). Scale bar=200 µm. a, b, c, d, and e represent higher magnification of the NTS in A, B, C, D, and E (Scale bar=100 µm).
|
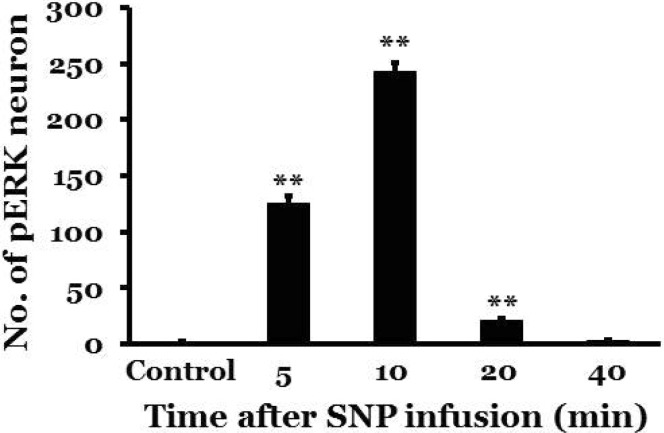 | Fig. 3Temporal changes in pERK expression in the nucleus tractus solitarius following a 30% reduction in blood pressure that was induced by SNP in conscious rats with intact labyrinths and baroreceptors. Controls received intravenous administration of saline at the same volume as SNP; min indicates the number of minutes after SNP administration. There were 8 rats in each group. *indicates a significant difference from the control group (**p<0.01).
|
Effect of BL or SAD on expression of pERK-immunoreactive neurons following acute hypotension
In rats with BL and/or SAD, a few pERK-immunoreactive neurons appeared in the NTS following saline infusion. However, expression of pERK-immunoreactive neurons in the sham group, in which the sinoaortic baroreceptors and vestibular end organs were intact, markedly increased in the NTS 10 min after administration of SNP (249.5±7.6), which was similar to levels observed in control rats with intact labyrinths and baroreceptors. In the BL group, expression of pERK-immunoreactive neurons increased in the NTS 10 min after administration of SNP (57.0±4.3), but the number of pERK-immunoreactive neurons was reduced by approximately 77.2% compared to the sham group. The expression was observed mainly in the caudal portion of the NTS. In the SAD group, increased expression of pERK-immunoreactive neurons was observed in the NTS 10 min after administration of SNP (150.8±4.9), which represents an approximate 39.6% reduction compared to the sham group. The expression of pERK-immunoreactive neurons was mainly located in the caudal portion of the NTS. In the BL+SAD group, SNP-induced acute hypotension did not affect the expression of pERK-immunoreactive neurons in the NTS (2.4±0.8), and the expression level was similar to that of control animals (Fig. 4).
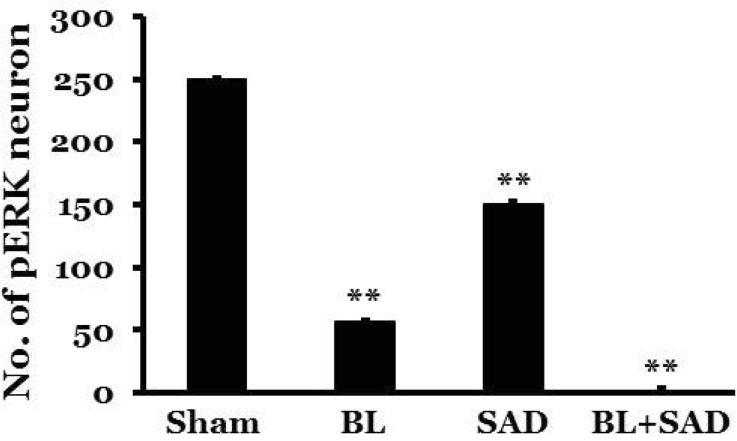 | Fig. 4Effect of acute hypotension on pERK expression in the nucleus tractus solitarius following a 30% reduction in blood pressure that was induced by SNP infusion in conscious rats with bilateral labyrinthectomy (BL), sinoaortic denervation (SAD), or both bilateral labyrinthectomy and sinoaortic denervation (BL+SAD). Expression of pERK was measured 10 min after SNP infusion. Sham, sham-treated. There were 8 rats in each group. *indicates a significant difference from the sham group (**p<0.01).
|
Go to :
DISCUSSION
Patients with idiopathic hypertension, myocardial infarction, arrhythmia, or congestive heart failure complain of vertigo, which could be explained by changes in blood flow to the peripheral vestibular system [1,23]. Animals with BL have reduced blood pressure when their body posture changes from a lying to a standing position [9], and patients with bilateral vestibular loss often complain of orthostatic hypotension [24]. These reports indicate that the vestibular system plays an important role in maintaining blood pressure during postural changes.
The NTS receives primary input from cardiovascular, gastrointestinal, and pulmonary afferents, and is a key component of the central pathway controlling sympathetic vasomotor activity [25]. Activation of baroreceptors showed a population of c-Fos protein expression situated at the dorsal region of the commissural part of the NTS and extending rostrally into circumscribed regions of the dorsal subnucleus [26], which is similar to the pattern of the central projections of the carotid sinus nerve and aortic depressor nerve. Distribution of vestibular projections was reported in the NTS; the caudal and intermediate portions of the NTS receive projections from the medial and inferior vestibular nuclei in rabbits [10], and the middle and lateral region of the NTS receive inputs from the medial and inferior vestibular nuclei in cats [11]. Thus, some neurons with vestibular inputs in the NTS receive convergent signals from the baroreceptors.
SNP decreases blood pressure by inducing the release of nitric oxide [27]. Several studies have reported that SNP-induced hypotension increases c-Fos protein expression in the supraoptic nucleus, paraventricular nucleus, rostral ventrolateral medullary nucleus, NTS, and vestibular nucleus [18,28]. In this study, acute hypotension induced by SNP infusion increased pERK expression in the NTS, which was reduced by removal of the baroreceptor or vestibular receptor inputs. According to our previous studies, acute hypotension induced by hemorrhage or SNP infusion increases neuronal activities [29] and expression of c-Fos protein [18] and pERK [19] in the medial vestibular nucleus, and these effects are abolished by the loss of peripheral vestibular receptors. Excitation in the vestibular nucleus might partly result from ischemic activation, excitotoxicity, of the hair cells from a reduction in blood flow to the inner ear following acute hypotension [29]. Our previous studies indicated that acute hypotension increases glutamate concentration in the vestibular nuclei [30,31,32]. Therefore, acute hypotension activates the vestibular receptors, afferent signals from the vestibular receptors project to the NTS through the vestibular nuclei, and the NTS integrates afferent signals from the baroreceptors and vestibular receptors. Expression of pERK in the NTS following acute hypotension resulted from activation of the NTS neurons through the vestibular receptor and baroreceptor inputs. These results are consistent with pERK expression resulting from activation of the NTS neurons following hydralazine induced hypotension [33].
Minimal pERK expression in the NTS was observed in the saline control group, indicating that the NTS did not receive any significant afferent signals in conscious animals during the experiment. The number of neurons expressing pERK following acute hypotension was reduced by approximately 39.6%, 77.2%, and 99.0% in the SAD, BL, and BL+SAD groups, respectively, compared to the sham group. In addition, 59.4% of neurons expressing pERK in the NTS received projections from the vestibular receptors, and 21.8% of neurons expressing pERK received projections from the baroreceptors. Therefore, expression of pERK in conscious animals with acute hypotension is largely a consequence of activation of the vestibular receptors. pERK expression in the NTS following acute hypotension in BL+SAD animals was reduced by approximately 99%, which indicates that pERK expression in the NTS following acute hypotension is likely due to both baroreceptor and vestibular receptor inputs. Expression of pERK following acute hypotension was mainly observed in the caudal portion of the NTS in BL and SAD animals, which is the same expression pattern previously observed for c-Fos protein following acute hypotension [34]. These results indicate that the main convergence area of the afferent inputs from the vestibular receptor and baroreceptor following acute hypotension might be the caudal portion of the NTS. Thus, the vestibular receptor and baroreceptor play a significant role in the regulation of blood pressure following acute hypotension; in particular, the vestibular system is considered to play a key role in compensating for orthostatic hypotension [35].
Expression of pERK in the NTS following acute hypotension observed in this study is not completely consistent with expression of c-Fos protein observed in our previous study [34]. The expression of c-Fos protein in the NTS following acute hypotension was greatly reduced by SAD and only moderately reduced by BL, and the number of neurons expressing c-Fos protein was reduced relative to those expressing pERK in the NTS following acute hypotension. Since pERK is a transcription factor of immediate early genes including c-fos [16], the numbers of neurons expressing c-Fos protein and pERK are not expected to be equivalent since the time required for pERK expression is much shorter compared to that for c-Fos protein. Both c-Fos protein and pERK can be used as markers for neuronal activation following several kinds of stimulation, but pERK is much more dynamic and appears to be a better marker for central sensitization [36]. Further studies will address other factors to explain the difference between the expression of c-Fos protein and pERK in the NTS following acute hypotension.
In summary, expression of pERK in the NTS induced by acute hypotension occurs in response to signals originating from the vestibular receptors and baroreceptors in conscious rats. In particular, the vestibular system is considered to play a key role in compensating for orthostatic hypotension. pERK expression in the NTS was localized to the caudal portion of the nuclei in the BL and SAD groups.
Go to :
 XML Download
XML Download