

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Recent large-scale epidemiological studies have revealed that major depression and anxiety are the most common psychiatric disorders in humans [1,2]. Although the etiology of depression- and anxiety-related disorders is still incompletely understood, stress is a potent environmental risk factor for these disorders. Chronic exposure to stress induces modifications in brain physiology and neural circuits. Stress triggers the rapid activation of the hypothalamic-pituitary-adrenal (HPA) axis in animals, leading to the release of corticosteroids from the adrenal cortex [3,4,5]. Corticosteroids pass the blood brain barrier and reach various brain regions via the circulation. Binding of corticosteroids to two related receptor molecules, the glucocorticoid receptor and the mineralocorticoid receptor, induces the following structural and functional changes in brain neurons: regression of dendritic processes, loss and shrinkage of dendritic spines, inhibition of neurogenesis, disturbance of synaptic plasticity, and neurodegeneration [6,7,8]. These deleterious effects of corticosteroids on neurons are believed to cause depression- and anxiety-related behavioral changes in animals. In agreement with these observations, the hyperactivity of the HPA axis, hypercortisolemia, or prolonged glucocorticoid administration is associated with depression- and anxiety-related disorders [9,10]. In addition, depression- and/or anxiety-like behaviors can be induced in rodents by various chronic stress paradigms in which animals are exposed to several weeks of repeated physical stressors or social stressors.
Interestingly, however, some chronic stress paradigms induce both anxiety and depression-like behaviors in animals, while others have been reported to increase one specific phenotype [11,12,13,14,15,16]. Considering the high comorbidity of depression and anxiety in humans, the coexistence of these disorders in rodents also seems possible. However, there is a possibility that the different behavioral outcomes among the previous studies were caused by different stress paradigms. This idea raises the question whether distinct types of stressors induce different behavioral changes. A few recent reports on social stress paradigms provide evidence supporting this hypothesis. For example, mice exposed to social defeat or chronic subordinate colony housing displayed increased anxiety behaviors without changes in depression-related behaviors [12,13,14]. In addition, social isolation also produced similar behavioral changes in mice [17]. These findings have led us to examine whether chronic stress without social factors selectively induces depression-like behaviors or both depression- and anxiety-like behaviors in mice. In the present study, we investigated the effects of three non-social stress paradigms on depression- and anxiety-related behaviors. Our study provides experimental evidence that social and non-social chronic stressors differentially affect depression and anxiety. Although psychosocial stress is more prevalent in humans, the dissociation of critical risk factors for depression and anxiety may provide clues to understand the neural mechanisms of psychiatric disorders.
METHODS
Animals
Four-week-old male C57BL/6N mice were purchased from Orient Bio (Gyeonggi, Korea). Upon arrival, the animals were acclimated for 1 week in our specific pathogen-free barrier facility before the experiments began. During the rearing and behavioral experiments, mice were housed in groups of 3~4/cage and monitored daily for signs of bullying, hair loss, or fight wounds. Except for stress sessions, animals were allowed free access to food and water and were maintained on a 12-hour light-dark cycle (lights on 08:00 h) at 23±1℃ with a relative humidity of 50~60%. The maintenance of all animals and related experiments were performed according to institutional guidelines for the care and use of animals in research (SNU-120323-1).
Experimental design for behavioral experiments
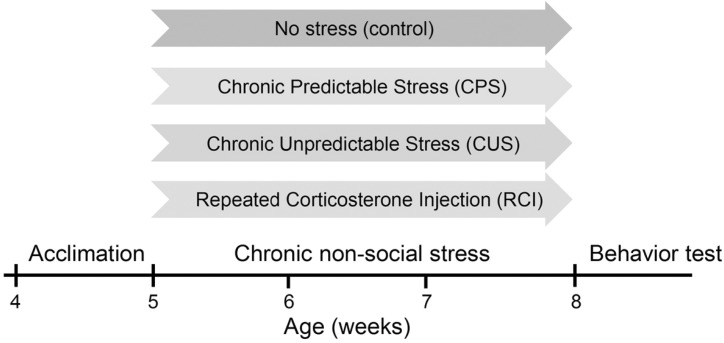
To investigate the effect of chronic non-social stress on depression- and anxiety-like behaviors in mice, male littermates were assigned to experimental or control groups at 35~38 days after birth. Each experimental group was exposed to a different type of chronic non-social stress for 3 weeks, as illustrated in Fig. 1. All of the three stress paradigms have been reported to cause stress responses, including increased corticosterone levels, in rodents [18,19,20,21,22]. For chronic predictable stress (CPS), mice were placed in a well-ventilated 60-ml syringe for 2 hours once per day between 17:00 and 19:00. During the 21 restraint sessions, mice were able to move laterally, but not vertically. For chronic unpredictable stress (CUS), mice were exposed in a random order to a variety of chronic stressors, including a wet cage (12 h), light-dark cycle reversal (24 h), white noise (100 dB, 12 h), cold water swim (10℃, 1 h), restraint (2 h), cage shake (30 rpm, 12 h), and electric foot shocks (10 scrambled shocks with duration of 2 s over 120 min). Animals were subjected to one stressor daily for 21 days. For repeated corticosterone injection (RCI), corticosterone-HBC complex was purchased from Sigma-Aldrich (St. Louis, MO) and dissolved in physiological saline. Mice received subcutaneous injections of the corticosterone-HBC complex (20 mg/kg) once per day for 21 days. The control animals for the repeated corticosterone-HBC injections were administered vehicle. All solutions were sterilized through a 0.22-µm filter before injection.
Open field test
Each animal was placed in the center of an open field apparatus with opaque walls (40×40×40 cm) in a dimly lit room. The behavior of each mouse was monitored for 30 minutes by video recording. The total distance traveled and time spent in the entire open field and in the center (20×20 cm) were calculated using video tracking software (Ethovision XT, Noldus, Netherlands). To avoid possible abnormal behaviors caused by the tail suspension test, the open field test was performed 1 day before the tail suspension test.
Tail suspension test
Mice were suspended by their tails from a steel bar using adhesive tape in a chamber with opaque walls. The distance between the floor of the chamber and the steel bar was approximately 40 cm. Mice that climbed onto their tail or fell off during the test were excluded from analysis. The total duration of immobility over the 6 min observation period was scored by an experimenter blinded to the experimental details.
Statistical analysis
Data were analyzed using IGOR Pro (WaveMetrics, OR, USA) and SPSS (Statistical Package for the Social Sciences, IBM, NY, USA) software. Statistical significance was determined by parametric two-tailed Student's t-tests and nonparametric Mann-Whitney-Wilcoxon tests. All data and error bars are expressed as the mean±s.e.m.
RESULTS
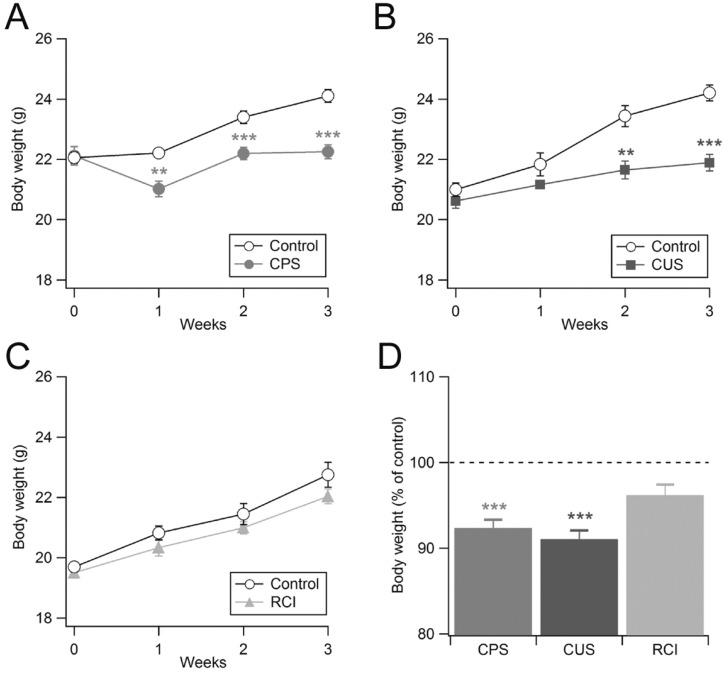
Effect of chronic non-social stress on body weight
Because weight changes have been observed in rodent models of chronic stress [23,24,25,26], we measured the daily body weight of mice during the stress period. As shown in Fig. 2A, CPS significantly reduced body weight. Repeated restraint caused weight loss during the first week and inhibited body weight gain through the end of the stress paradigm. After 3 weeks, stressed animals gained 0.7% of body weight (22.12±0.30 g and 22.25±0.23 g at weeks 0 and 3, respectively; n=9), whereas the body weight of control mice increased 9.3% (22.05±0.19 g and 24.10±0.21 g at weeks 0 and 3, respectively; n=9). These results suggest a significant interaction between body weight gain and treatment (p<0.001, Student's t-test and Mann-Whitney-Wilcoxon test).
To investigate the relationship between the predictability of stress and the reduction of body weight, we next examined the effect of chronic unpredictable non-social stress on body weight (Fig. 2B). Similar to CPS, weight gain during the 3 weeks was greater in the control animals (15.2%; 21.0±0.66 g and 24.20±0.78 g at weeks 0 and 3, respectively; n=9) than in the stressed animals (6.2%; 20.63±0.68 g and 21.88±0.77 g at weeks 0 and 3, respectively; n=8). This result indicates that CUS reduces body weight gain in mice and that the predictability of stress is not important for its effect on body weight gain (CPS: 92.34±0.96% of control; CUS: 91.05±1.01% of control; p>0.1; Fig. 2D).
We next examined the effect of predictable non-physical stress on body weight. Mice that received RCI showed comparable body weight gain to control mice that received repeated injections of vehicle (Fig. 2C). The control mice (n=10, 19.69±0.52 g and 22.75±1.30 g at weeks 0 and 3, respectively) and the corticosterone-HBC complex-treated mice (n=11, 19.35±0.57 g and 22.16±0.85 g at weeks 0 and 3, respectively) gained 15.5% and 14.5% of their original body weight, respectively. This difference did not reach statistical significance (p>0.1). Therefore, in contrast with CPS or CUS, RCI had no effect on body weight gain (Fig. 2D).
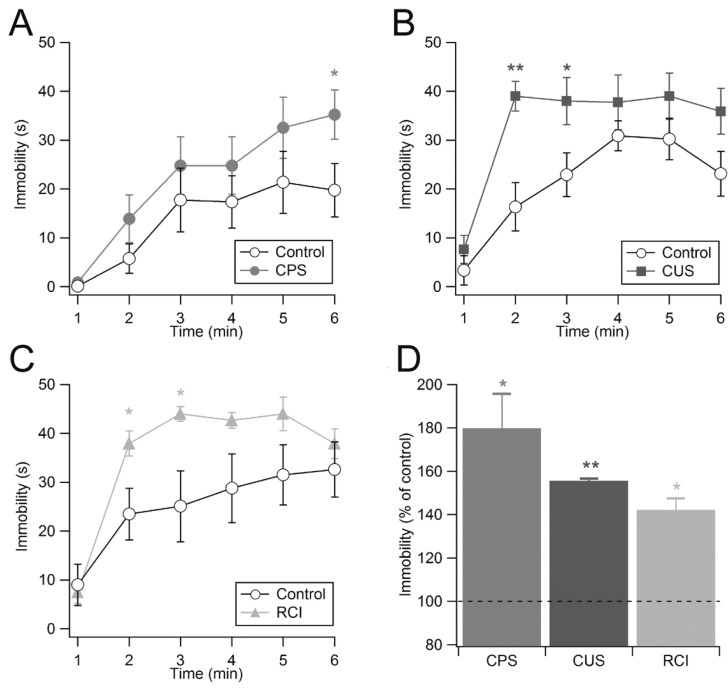
Effect of chronic non-social stress on depression-like behaviors
To investigate depression-like behaviors, we assessed immobility/despair behaviors of mice using the tail-suspension test [27]. In this test, immobility is thought to reflect a state of despair and of giving up trying to escape from an uncomfortable situation.
Mice exposed to CPS displayed high levels of immobility in the tail-suspension test (Fig. 3A). During the 6-minute test, the total duration of immobility in the control group and in the stressed group was 82.1±18.7 s and 147.63±13.1 s, respectively (n=8 pairs, p<0.05; Fig. 3D). We next examined the effect of CUS on depression-like behaviors. Similar to repeated restraint, the mice exposed to CUS spent more time immobile compared to the control mice (control: n=9, 126.7±10.4 s; CUS: n=8, 197.3±15.2 s; p<0.01; Fig. 3B, 3D). Consistently, a minute-by-minute analysis of the time spent immobile revealed that the CUS group became immobile significantly sooner than the control mice (Fig. 3B). We further tested RCI, predictable non-physical stress, on depression-like behaviors. As shown in Fig. 3C and Fig. 3D, RCI significantly increased the amount of time mice remained immobile by 42.1% compared to the vehicle-treated control mice (control: n=10, 150.5±29.9 s; RCI: n=11, 213.9±26.4 s; p<0.05; Fig. 3C, 3D). Together, these results suggest that each of the three chronic non-social stress paradigms caused elevated levels of depression-like behaviors in the tail-suspension test, regardless of the predictability.
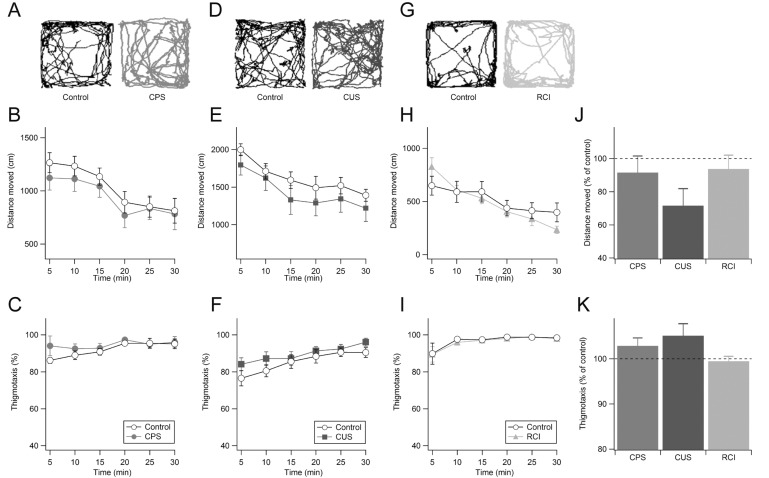
Effect of chronic non-social stress on anxiety-related behaviors
To test whether chronic non-social stress induces both depression- and anxiety-related phenotypes, we examined anxiety-related behaviors using the open field test [28,29]. In this test, the degree of thigmotaxis, the tendency to remain close to the walls, is considered an index of anxiety in mice [30]. Because anxious animals are less likely to explore new environments, they show enhanced thigmotaxis and reduced locomotor activity in the open field chamber. We analyzed thigmotaxis and locomotor activity by measuring the time spent in the peripheral areas and the distance moved in the open field chamber, respectively.
Mice exposed to CPS displayed normal locomotor activity and thigmotaxis levels compared to their control littermates during all test periods (Fig. 4A-C). In addition, when we analyzed the total distance moved (control: n=9, 61.90±4.96 m; CPS: n=9, 56.67±6.22 m; p>0.1; Fig. 4J) and the time spent in the peripheral area (1656.0±37.9 s vs. 1703.0±29.4 s; p>0.1; Fig. 4K) during the 30-minute test periods, we could not find any differences between the two groups. We next examined CUS on anxiety-related behaviors (Fig. 4D). Although mice exposed to CUS showed a tendency to be hypoactive (Fig. 4E) and enhanced thigmotaxis (Fig. 4F), the difference did not reach statistical significance during any test period. When we analyzed the whole test session, we also found no significant effects of CUS on total distance moved (control: n=9, 97.10±5.21 m; CUS: n=8, 88.47±9.03 m; p>0.1; Fig. 4J) and thigmotaxis (1536.0±43.2 s vs. 1614.7±40.6 s; p>0.1; Fig. 4K). Additionally, we examined the effects of RCI on anxiety-related behaviors (Fig. 4G). Again, we did not detect any significant differences in distance moved (Fig. 4H) and time spent in the peripheral areas (Fig. 4I) between the corticosterone- and vehicle-treated groups. As shown in Fig. 4J and Fig. 4K, both groups of mice showed similar levels of exploratory activity (control: n=10, 31.39±4.12 m; RCI: n=11, 29.42±2.62 m; p>0.1) and thigmotaxis (control: 1741.8±18.7 s; RCI: n=11, 1732.3±18.7 m; p>0.1) during the entire test session.
Taken together, these results indicate that each of the three chronic non-social stress paradigms had no effect on anxiety-related behaviors of mice.
DISCUSSION
Although stress is a risk factor for depression- and anxiety-related disorders, the differential effects of social and non-social stress on these neuropsychiatric disorders are poorly understood. In the present study, we examined the effect of three chronic non-social stress paradigms on mouse behavior. We found that CPS, CUS, and RCI induced depression/despair-like behaviors without affecting anxiety-related behaviors.
According to human studies [31,32,33,34], depression and anxiety show high comorbidity rates. Approximately half of patients with major depressive disorder met the criteria for generalized anxiety disorder, and the majority of patients with generalized anxiety disorder also have major depressive disorder [33,35]. These findings imply that brain circuits associated with depression partly overlap with those of anxiety disorder and that certain types of stress can modify brain circuits associated with both disorders [5,36]. Although the exact brain circuits that mediate depression and anxiety are still unclear, recent human imaging studies identified some brain areas dysregulated in depression and anxiety disorders, including the prefrontal cortex, subgenual cingulate cortex, hippocampus, and amygdala [37,38]. These brain areas are implicated in the regulation of stress responses as well as emotional functions [39]. In mammals, stress activates the HPA axis and results in the secretion of stress hormones (corticosteroids) from the adrenal cortex. Although the neural mechanism that translates stressful stimuli into the activation of the paraventricular nucleus (PVN) in the hypothalamus is still incompletely understood, the hippocampus appears to inhibit the PVN through the ventral subicular projection to PVN-connected GABAergic neurons in the bed nucleus of the stria terminalis (BNST) [5]. In contrast, the amygdala appears to activate hypothalamic areas through excitatory and double GABAergic projections to the PVN [4]. In accordance with this observation, hippocampal stimulation decreases plasma corticosteroids in rats and humans, while stimulation of the amygdala produces the opposite effect [40,41]. Furthermore, overactivation of the amygdala is associated with depression and anxiety [37,38].
In the present study, the three non-social stress paradigms induced a reduction in body weight gain and increased immobility in the tail-suspension test (Fig. 2 and 3). Although decreased weight gain under stress does not necessarily reflect the loss of appetite in animals, the loss of interest in enjoyable activities (anhedonia) is one of the core symptoms of major depressive disorder. In addition, despair (feelings of hopelessness) is the most common symptom of depression. Our study indicates that depression- and anxiety-related circuits modified by stress can be dissociated in the mouse brain, and distinct types of stress affect different brain circuits.
There is a possibility that social stress modifies brain circuits related to anxiety as well as depression, while non-social stress influences depression-related brain circuits. Interestingly, psychosocial stress rather than non-social stress seems more relevant in humans. Thus, the social component of stress might be one of the possible explanations for the high rate of comorbidity between anxiety and depression in human patients. Consistent with this idea, several recent lines of evidence suggest a strong association between social stress and anxiety in rodents. Social isolation, a mild social stress paradigm, selectively induced anxiety-related behavioral changes in mice without affecting depression-like behaviors [17]. In addition, repeated social defeat and chronic subordinate colony housing caused increased anxiety-like behavior in mice, but depression-like behaviors were not changed by these social stress paradigms [12,13,14,15]. However, some experimental evidence argues for a selective effect of chronic social stress on anxiety [11,14,16]. Interestingly, a significant interaction between behavioral changes and time of stress was observed. Stressed animals displayed reduced exploratory activity one week after stress, but anhedonia could be observed 3 weeks after chronic social stress [11]. These results imply that prolonged social stress (more than 3 weeks) might induce depression- as well as anxiety-related behavioral changes in rodents [14,16]. Whether prolonged (more than 4 weeks) non-social stress influences anxiety-related brain circuits and induces anxiety-related behavioral changes as well as depression deserves further investigation.
 XML Download
XML Download