

 PDF
PDF ePub
ePub Citation
Citation Print
Print


ABBREVIATIONS
EFA
essential fatty acid
LA
linoleic acid
ALA
α-linolenic acid
OA
oleic acid
PO
palmitic acid
POA
palmitoleic acid
araA
adenine 9-β-D-arabinofuranoside
2-NBDG
2-[N-(7-nitrobenz-2-oxa-1,3-diazol-4-yl)amino]-2-deoxy-D-glucose
MTT
3-(4,5-Dimethyl-2-thiazolyl)-2,5-diphenyltetrazolium bromide
PKCθ
protein kinase Cθ
JNK
Jun-N-terminal kinase
AMPK
AMP-activated protein kinase
ACC
acetyl-CoA carboxylase
INTRODUCTION
Skeletal muscle is a primary site of glucose utilization in the human body. First of all, insulin stimulates glucose uptake through the complex insulin signaling pathways in skeletal muscle cells. Binding of insulin to its receptors causes phosphorylation of insulin receptor substrate-1 (IRS-1) that then engages and activates phosphatidylinositide 3-kinase (PI3K). This induces the sequential activation of Akt and Akt substrate-160 kDa (AS160), which results in translocation of the glucose transporters to the plasma membrane from its intracellular pool and then transports glucose into the cells [1,2]. Meanwhile, phosphorylation of certain serine residues on IRS-1 usually inhibits this insulin signaling. Phosphorylation of these serine residues interrupts the normal interaction between IRS-1 and insulin receptor, thereby blocking the downstream spread of insulin signaling. Some serine/threonine kinases, including PKCθ, JNK, and IκB kinase, are involved in the inhibitory phosphorylation of IRS-1 on serine residues [3,4]. It has been suggested that the excess uptake of saturated fatty acid by the myocyte results in the cellular accumulation of intermediates such as diacylglycerol and ceramide, which causes the activation of serine/threonine kinases and the sequential inhibition of IRS-1/PI3K/Akt pathway and then ultimately induces insulin resistance [5,6,7]. The saturated fatty acids such as palmitic acid (PA) have been reported to induce insulin resistance and apoptosis, whereas monounsaturated fatty acid such as oleic acid (OA) protects cells from deleterious effects of PA [5,7,8,9,10]. It has been recently reported that OA reverses PA-induced insulin resistance by promoting triglyceride accumulation and mitochondrial β-oxidation in C2C12 skeletal muscle cells [7] and maintaining insulin signaling through PI3K in L6 skeletal muscle cells [8].
On the other hand, a metabolic master switch in the cells, AMP-activated protein kinase (AMPK) is another important signaling molecule that promotes intracellular glucose uptake independently from insulin. The regulation of AMPK activity in skeletal muscle is coordinated by upstream kinases, the intracellular ratio of AMP:ATP, and a growing number of extracellular factors such as hormones, cytokines, nutrients, and exercise [11,12]. Increased glucose uptake and fatty acid oxidation in response to AMPK activation may occur via phosphorylation of downstream Rab GTPase-activating proteins and acetyl-CoA carboxylase (ACC) respectively [11,13].
Essential fatty acid (EFA) is a polyunsaturated fatty acid that humans must incept from nourishment because EFA is essentially required to maintain good health but not synthesized in the body. EFA plays roles in the regulation of many metabolic processes in the cells. It has been suggested by many investigators that deficiency of EFA or the wrong balance of EFAs may be a risk factor for many metabolic syndromes such as cardiovascular diseases and osteoporosis [14,15]. Humans cannot form the double bonds in fatty acids beyond carbon 9 and 10, so ω-6 linoleic acid (LA, C18:2) and ω-3 α-linolenic acid (ALA, C18:3) are essential for humans in their diet. Conditionally essential long-chain polyunsaturated fatty acids such as γ-linolenic acid (C18:3), arachidonic acid (C20:4), eicosapentaenoic acid (C20:5), and docosahexaenoic acid (C22:6) are known to be synthesized from LA and ALA in humans.
Several unsaturated fatty acids were reported to affect glucose uptake in skeletal muscle cells. One of the most common monounsaturated fatty acids, OA, has been reported to protect against PA-induced impairment in glucose uptake and insulin signaling through PI3K activation and up-regulation of insulin receptor expression [8,16]. It has recently published that several polyunsaturated fatty acids including EFAs ameliorate PA-induced impairment of 2-deoxy glucose uptake in L6 muscle cells [16]. However, cellular mechanisms underlying this protective effects of EFAs against the deleterious action of PA have not yet been determined up to the present time. On the other hand, relatively many studies have been carried out to examine the effects of conjugated linoleic acids (CLAs), which are structural isomers of linoleic acid, on myocellular glucose uptake because natural CLAs are mainly present in ruminant products such as meat, milk and dairy products. It has been reported that AMPK signaling pathway (Ca2+/calmodulin-dependent protein kinase II-AMPK-AS160) is involved in the regulation of glucose uptake by cis-9, trans-11-CLA and trans-10, cis-12 CLA in skeletal muscle cells [17,18,19]. In this study, we examined the effect of EFA on glucose uptake in C2C12 skeletal muscle cells and investigated possible cellular mechanisms underlying EFA's actions.
Go to : 
METHODS
Materials
Mouse C2C12 skeletal muscle cells were obtained from the ATCC (Rockville, USA). Fetal bovine serum (FBS), horse serum, Dulbecco's modified eagle medium (DMEM), no-glucose DMEM, Dulbecco's phosphate buffered saline (DPBS), and Antibiotic-Antimycotic were from Gibco (Grand Island, USA). Fatty acid free-bovine serum albumin (BSA) was obtained from Fitzgerald (North Acton, USA). Palmitic acid (PA), palmitoleic acid (POA), oleic acid (OA), linoleic acid (LA), α-linolenic acid (ALA), bovine insulin, metformin, and adenine 9-β-D-arabinofuranoside (araA) were obtained from Sigma-Aldrich (St. Louis, USA). 2-[N-(7-nitrobenz-2-oxa-1,3-diazol-4-yl)amino]-2-deoxy-D-glucose (2-NBDG) was obtained from Invitrogen (Carlsbad, USA). 3-(4,5-Dimethyl-2-thiazolyl)-2,5-diphenyltetrazolium bromide (MTT) was obtained from Amresco (Solon, USA). The antibodies specific for phospho-Akt (Ser473), Akt, phospho-PKCθ (Thr538), PKCθ, phospho-JNK (Thr183/Tyr185), JNK, phospho-AMPKα (Thr172), AMPKα, phospho-acetyl-CoA carboxylase (ACC) (Ser79), and ACC were obtained from Cell Signaling Technology (Danvers, USA).
Cell culture and fatty acid treatment
C2C12 cells were cultured in DMEM containing 10% FBS, 100 units/ml penicillin, 100 µg/ml streptomycin, and 250 ng/ml amphotericin B at 37℃ under a humidified condition of 95% air and 5% CO2. When cells reached confluence, the medium was replaced by differentiation medium (DMEM containing 2% horse serum) to induce differentiation from myoblasts into fused myotubes as a previously described method [5]. The differentiation medium was changed every other day. After 4 days, experiments were conducted. To induce insulin resistance in the differentiated C2C12 cells, the medium was replaced by a medium containing PA, which was prepared by the conjugation of PA with fatty acid (FA)-free bovine serum albumin (BSA) as a previously described method with slight modifications [9,10]. Briefly, PA was dissolved in absolute ethanol at a concentration of 100 mM and diluted to a ratio of 1:200 with DMEM containing 2% FA-free BSA, and then incubated with shaking for 3 h at 37℃ prior to the experiment initiation. Differentiated C2C12 cells were routinely incubated for 18 h in the above medium in the presence of 500 µM PA either alone or in combination with other FA. Different concentrations of BSA-conjugated FA were also prepared by the same method by the conjugation with 10~100 mM FA (POA, OA, LA, or ALA) with the appropriate concentration of FA-free BSA (0.2~2% BSA). The molar ratio of FA/BSA in this preparation is similar to that observed in human serum [20].
Measurement of glucose uptake activity using 2-NBDG
Glucose uptake activity was measured using a fluorescent D-glucose analogue 2-[N-(7-nitrobenz-2-oxa-1,3-diazol-4-yl)amino]-2-deoxy-D-glucose (2-NBDG) in C2C12 cells as a previously described method with slight modifications [21,22]. Briefly, differentiated muscle cells on 6-well plates were treated with different kinds of BSA-conjugated FA for 18 h. To determine the involvement of AMPK in EFA-stimulated glucose uptake, 2 mM araA (a competitive inhibitor of AMPK) was added to a medium at 20 min before the incubation with FA. After the incubation, cells were washed with DPBS and incubated with 100 nM insulin in no-glucose DMEM for 10 min and 60 µM 2-NBDG was added to a medium for another 1 h. The medium was then washed twice with cold DPBS to remove free 2-NBDG. The cells in each well were suspended with DPBS after trypsinization and subsequently transferred to 96 black well fluorescence plates. The fluorescence intensity of cellular 2-NBDG in each well was measured at an excitation wave-length of 485 nm and an emission wavelength of 535 nm using Fluorescent microplate reader (Molecular Devices, USA).
Cell viability assay
The cell viability was measured using a MTT-based colorimetric assay as a previously described method with slight modifications [23]. Briefly, after the treatment of BSA-conjugated FA, cultured cells in 24-well plates were washed with DPBS and incubated with phenol red-free DMEM containing 1 mg/ml MTT for 3 h at 37℃, and then 0.5 ml 2-propanol was added to each well for 30 min. The amount of dissolved MTT-derived formazan was estimated by spectrophotometer to assess the living cells in culture. Data were displayed as a percentage of untreated control.
Western blot analysis
The whole cell lysate was prepared by washing the cells twice with cold DPBS followed by the treatment of lysis buffer (150 mM NaCl, 50 mM Tris, 1 mM phenylmethanesulfonyl fluoride, 1 mM Na3VO4, 1% Nonidet P-40, 0.1% SDS, 0.5% deoxycholic acid, 1% protease inhibitor cocktail, pH 7.5) for 20 min on ice. Proteins in the cell lysate were separated by SDS-polyacrylamide gel electrophoresis and transferred onto PVDF membrane. The membrane was incubated with 5% non-fat dry milk or 5% BSA, and then incubated with antibody towards pAkt, Akt, pPKCθ, PKCθ, pJNK, JNK, pAMPK, AMPK, pACC, and ACC for overnight at 4℃, respectively. Subsequently, appropriate horseradish peroxidase-linked secondary antibodies (Cell Signaling, Danvers, USA) were added. Immunoreactive band was exposed on X-ray film using the enhanced chemiluminescence western blotting detection system from GE Healthcare (Buckinghamshire, UK) according to the manufacturer's instruction. The density of each band was quantified using ImageQuant software. The level of each protein was normalized to that of control protein, β-actin.
Statistical analysis
All data in figures denote at least four independent experiments and are represented as means±SE. One-way ANOVA with post Dunnett's test was used for multiple comparison and two-tailed Student's t-test was used to compare values between two groups using the GraphPad Prism software. p-values p<0.05 were considered as statistically significant.
Go to :
RESULTS
Effect of fatty acid treatment on cell viability
In this study, glucose uptake activity was assessed in cells treated with fatty acids for 18 h, and hence it was necessary to determine whether fatty acid affects the cell survival of C2C12 cells. When C2C12 cells were incubated with different doses of palmitic acid (PA) for 18 h, cell viability decreased in a dose-dependent manner (Data not shown). Significant cell death was observed at 250 µM and 500 µM PA as compared with control survival. Meanwhile, oleic acid (OA), linoleic acid (LA), and α-linolenic acid (ALA) all did not affect the cell survival up to a concentration of 500 µM. In addition, PA-induced cytotoxicity was almost reversed by the co-treatment with the same concentration of OA, LA, and ALA, respectively (Data not shown). In this study, glucose uptake activity of C2C12 cells was calibrated by cellular uptake 2-NBDG (a fluorescent D-glucose analogue) intensity per cell viability in each well and displayed as a percentage of untreated control cells.
Effect of EFAs on PA-induced impairment in glucose uptake
2-NBDG uptake assay was used to assess the effects of essential fatty acids (EFAs), LA and ALA, on the PA-induced impairment of glucose uptake activity in C2C12 muscle cells. When cultured cells were incubated with different doses of PA for 18 h, 2-NBDG uptake gradually decreased in a dose-dependent manner as shown in Fig. 1A. Furthermore, insulin-stimulated glucose uptake was virtually dissipated in the cells treated with over 250 µM PA. These impairments in basal and insulin-stimulated glucose uptake were almost reversed by the co-treatment of the same concentration of EFAs or OA with 500 µM PA in culture media (Fig. 1B). However, there was no significant difference between EFAs and OA in terms of recuperating from PA-induced impairment of glucose uptake.
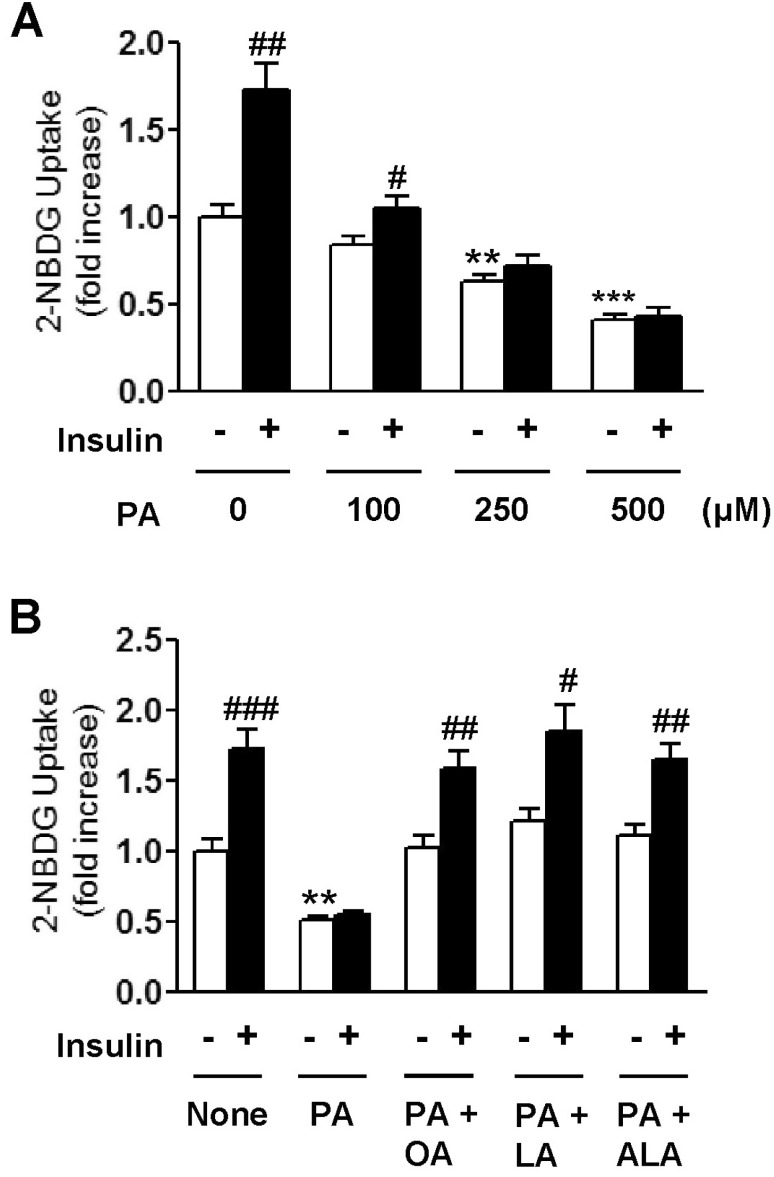 | Fig. 1Concentration-dependent effect of palmitic acid (PA) on 2-NBDG uptake of C2C12 cells in the presence or absence of 100 nM insulin (A). Effects of oleic acid (OA), linoleic acid (LA), and α-linolenic acid (ALA) on PA-induced impairment of 2-NBDG uptake in C2C12 cells (B). Cells were incubated with different kinds of 500 µM fatty acids either alone or in combination for 18 h. And then, cells were incubated with or without 100 nM insulin. Glucose uptake activity was measured using a fluorescent 2-NBDG as described in Methods. The results are presented as means±SEM of 6 independent experiments. **p<0.01, ***p<0.001 compared to untreated control group and #p<0.05, ##p<0.01, ###p<0.001 compared to insulin-untreated group.
|
Effect of EFAs on PA-induced suppression of insulin signaling
The most common saturated fatty acid, PA has been reported to impede normal insulin signal transduction and induce impaired glucose uptake in skeletal muscle. In this study, we examined whether polyunsaturated EFAs (LA; C18:2, ALA; C18:3) can reverse the PA-induced inhibition of Akt phosphorylation in comparison with monounsaturated OA (C18:1) and POA (C16:1). As shown in Fig. 2, the acute (10 min) treatment of 100 nM insulin induced a robust phosphorylation of Akt. However, this phosphorylation was significantly inhibited by the treatment of 500 µM PA, whereas unsaturated fatty acids (POA, OA, and EFAs) had no effect on the insulin-stimulated Akt phosphorylation (Fig. 2) as well as basal Akt phosphorylation (data not shown). The co-treatment of PA-exposed cells with increasing concentrations (50~500 µM) of C18 unsaturated fatty acids (OA and EFAs) reversed the PA-induced inhibition of Akt phosphorylation in a dose-dependent manner. However, C16 unsaturated fatty acid, POA, did not significantly reverse this inhibitory effect of PA, although POA also had a similar tendency to reduce the effect of PA on Akt activation.
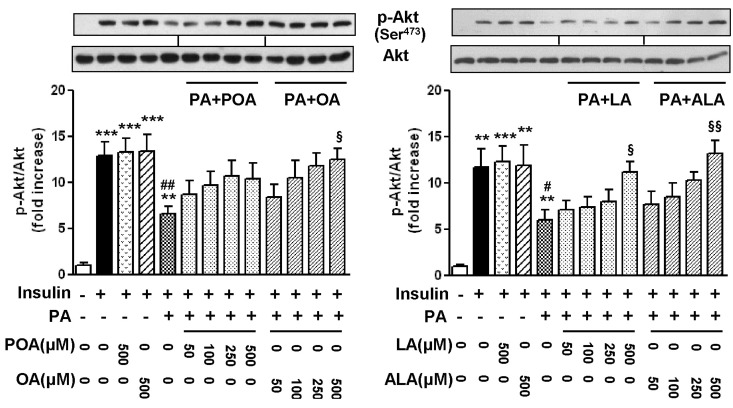 | Fig. 2Effects of palmitoleic acid (POA, C16:1), oleic acid (OA, C18:1), linoleic acid (LA, C18:2), and α-linolenic acid (ALA, C18:3) on the levels of phosphorylation of Akt in palmitic acid (PA)-treated C2C12 cells. Cells were incubated with different kinds and concentrations of fatty acids either alone or in combination for 18 h. And then, cells were incubated with or without 100 nM insulin. The level of p-Akt was detected by Western blotting. The results are presented as means±SEM of 4 independent experiments. **p<0.01, ***p<0.001 compared to untreated control group, #p<0.05, ##p<0.01 compared to insulin-treated group, and §p<0.05, §§p<0.01 compared to PA-treated group in the presence of 100 nM insulin.
|
It has been recently reported that PA induces insulin resistance by the activation of several serine/threonine kinases, including PKCθ, JNK, and IκB kinase [3,4]. Therefore, in this study, we subsequently examined whether EFAs can prevent the PA-induced activation of PKCθ and JNK. Consistent with previous studies by other investigators [3,4,5], 500 µM PA strongly stimulated PKCθ phosphorylation on Thr538, whereas the exposure to the same concentration of unsaturated fatty acid did not (Fig. 3A). Likewise, JNK phosphorylation on Thr183 and Tyr185 was also induced by the treatment of PA, whereas all of unsaturated fatty acids did not significantly affect the levels of phospho-JNK (Fig. 3B). The PA-induced phosphorylation of PKCθ and JNK were gradually inhibited by the co-treatment of PA-exposed cells with increasing concentrations of POA, OA, and EFAs, respectively. There was no distinguishable difference in potency inhibiting phosphorylation of serine/threonine kinases among four kinds of unsaturated fatty acids under test.
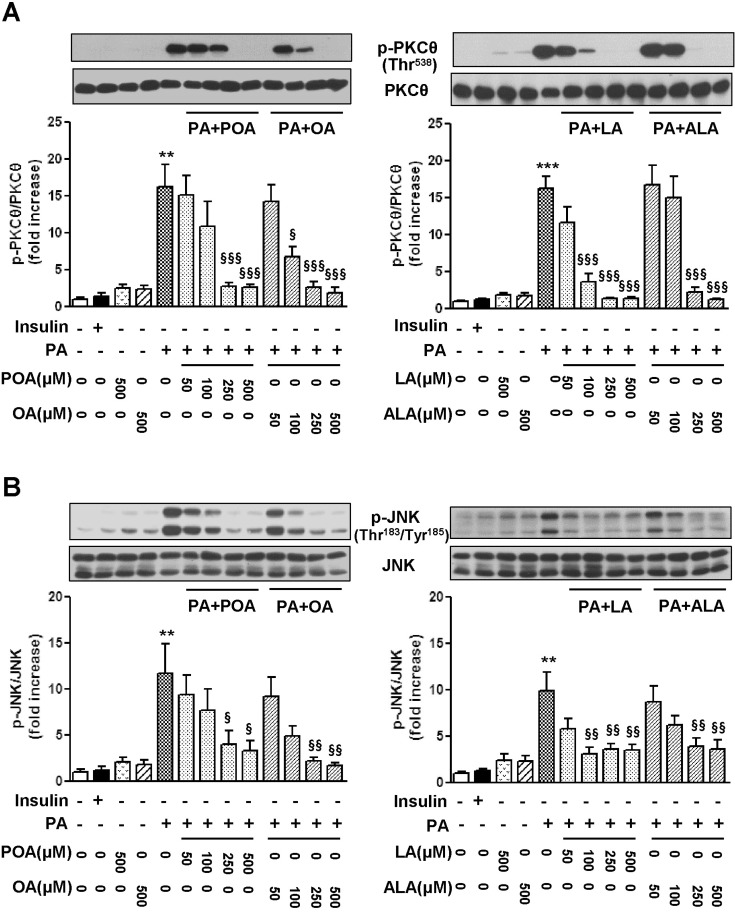 | Fig. 3Effects of palmitoleic acid (POA, C16:1), oleic acid (OA, C18:1), linoleic acid (LA, C18:2), and α-linolenic acid (ALA, C18:3) on the levels of phosphorylation of PKCθ (A) and JNK (B) in palmitic acid (PA)-treated C2C12 cells. Cells were incubated with different kinds and concentrations of fatty acids either alone or in combination for 18 h. The levels of p-PKCθ and p-JNK were detected by Western blotting. The results are presented as means±SEM of 4 independent experiments. **p<0.01, ***p<0.001 compared to untreated control group, and §p<0.05, §§p<0.01, §§§p<0.001, compared to PA-treated group.
|
Involvement of AMPK in glucose uptake stimulated by EFAs
We next investigated the effect of EFAs on glucose uptake at non-insulin resistant condition. As shown in Fig. 4A, when PA-untreated, control cells were incubated with 500 µM of EFAs (LA and ALA), OA and 1 mM metformin (an AMPK activator) for 18 h, respectively, 2-NBDG uptake was significantly increased by the treatment of EFAs and metformin. 2-NBDG uptake was also enhanced a little by OA, but not statistically significant and far weaker than by EFAs. To elucidate this difference in promoting glucose uptake between EFAs and OA, we examined whether EFAs can activate AMPK signaling pathway. The treatment with 500 µM EFAs and 1 mM metformin for 18 h caused significant phosphorylation of both AMPK and ACC, respectively (Fig. 4B-4D). However, OA did not significantly affect the levels of phosphor-AMPK and phosphor-ACC. We used adenine 9-β-D-arabinofuranoside (araA), a pharmacological inhibitor of AMPK, to confirm the involvement of AMPK pathway in promoting glucose uptake by EFAs. The pre-treatment of 2 mM araA did not affect levels of phospho-AMPK, phosphor-ACC, and 2-NBDG uptake. However, araA significantly abrogated the increase in the levels of AMPK and ACC phosphorylation as well as glucose uptake by EFAs.
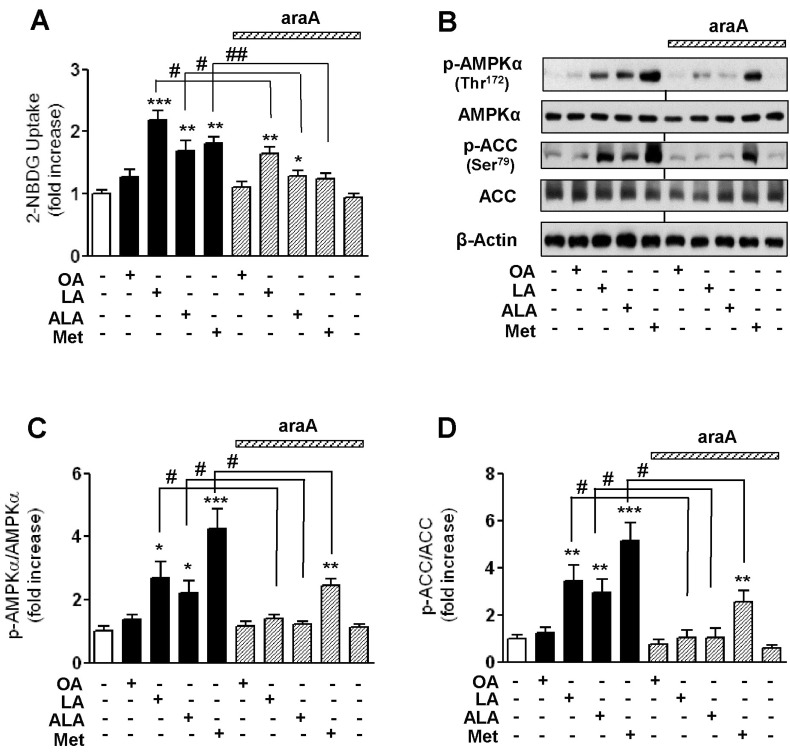 | Fig. 4Effects of oleic acid (OA), linoleic acid (LA), α-linolenic acid (ALA), and metformin (Met) on 2-NBDG uptake of C2C12 cells in the presence or absence of an AMPK inhibitor, araA (A) and the levels of phosphorylation of AMPK and ACC (B~D). Cells were incubated with different kinds of 500 µM fatty acids and 1 mM Met for 18 h. A specific AMPK inhibitor, araA (2 mM) was added to medium at 20 min before incubation of fatty acid. Glucose uptake activity was measured using a fluorescent 2-NBDG as described in Methods. The levels of p-AMPK and ACC were detected by Western blotting. The results are presented as means±SEM of 6 independent experiments. *p<0.05, **p<0.01, ***p<0.001 compared to untreated control group, #p<0.05, ##p<0.01 compared to araA-untreated corresponding group.
|
Go to :
DISCUSSION
Skeletal muscle is a primary site of glucose uptake and disposal after meal and, therefore, plays a crucial role in the maintenance of glucose homeostasis. Many investigators have reported that long-chain saturated fatty acids such as palmitic acid (PA) generally reduce insulin responsiveness and eventually cause impaired glucose uptake in skeletal muscle cells, whereas long-chain unsaturated fatty acids ameliorate these deleterious effects of saturated fatty acids [8,16,17,18,19]. However, the effect and the mechanism of polyunsaturated essential fatty acids (EFA) on glucose uptake have not yet been thoroughly investigated. In this study, we have demonstrated that two essential fatty acids (EFAs), linoleic acid (LA) and α-linolenic acid (ALA), stimulate glucose uptake through the restoration of suppressed insulin signaling pathway at PA-induced insulin resistant condition and the AMPK activation in C2C12 skeletal muscle cells. The C2C12 cells can be differentiated into fused myotubes from myoblasts in culture and therefore provide a useful model system for the study of cellular glucose uptake.
Consistent with previous studies by other investigators [16,22,24], we confirmed again that PA induced the impairment of both basal and insulin-stimulated glucose uptake of skeletal muscle cells in a dose-dependent manner (Fig. 1). We also found that these impairments in glucose uptake were nearly completely prevented by the treatment of the same concentration of EFAs and oleic acid (OA). However, there was no significant difference between EFAs and OA in terms of ameliorating PA-induced decrease in glucose uptake. This observation is consistent with a recently published investigation in which LA, ALA, and OA all similarly reversed a PA-induced decrease in a 2-deoxy glucose uptake in L6 myotubes [16]. Glucose uptake is occurred by two distinct cellular mechanisms in skeletal muscles. One is the insulin signaling pathway including the Akt activation, and the other is the insulin-independent AMPK pathway. Activated Akt and AMPK can increase the rate of the translocation of glucose transporter-4 stored in the cytosol to plasma membranes, causing enhancement of glucose uptake. Therefore, we assessed the effect of EFA on the activation of Akt and two serine/threonine kinases (PKCθ and JNK), which are involved in the inhibitory regulation of insulin signaling. Data showed that insulin-stimulated Akt phosphorylation was significantly decreased after the treatment of PA for 18h, whereas PKCθ and JNK phosphorylation were markedly induced by PA (Fig. 2 and 3). The effect of the PA treatment on the levels of Akt, PKCθ and JNK phosphorylation in skeletal muscle cells is consistent with findings reported previously [5,9,25]. In contrast, EFA and OA significantly inhibited PA-induced PKC and JNK phosphorylation and restored the level of Akt phosphorylation. The activation of PKCθ and JNK has been reported to induce serine phosphorylation of IRS-1 and inhibit an associated PI3K/Akt activity, thereby reducing the rate of glucose transport into the cells [3,4,5]. Therefore, the protective effect of EFA against the PA-induced reduction in glucose uptake could be due to the suppression of phosphorylation of these serine/threonine kinases. As compared the restoring potency of polyunsaturated EFAs (C18:2 and C18:3) in the PA-induced suppression of insulin signaling pathway with its corresponding monounsaturated OA (C18:1), there was no distinct difference between them, which is well correlated with the data obtained from the glucose uptake measurement in Fig 1. It has been reported that OA prevented PA-induced insulin resistance and inflammation in C2C12 cells by reducing the intracellular accumulation of diacylglycerol, which in turn activates the PKCθ [5]. However, it is still unclear whether EFA also affects the formation of lipid metabolites such as diacylglycerol or ceramide in PA-induced insulin resistant cells.
Next, we assessed glucose uptake activity by EFA at non-insulin resistant condition. As shown in Fig. 4, the incubation of PA-untreated, control cells with 500 µM EFA for 18 h caused a significant increase in glucose uptake. We also found that EFAs were more effective than OA in terms of promoting glucose uptake in control cells. Although OA slightly increased glucose uptake by C2C12 cells, but not statistically significant and the net increase was much smaller than those caused by EFAs and metformin. Metformin is a clinically used hypoglycemia drug, is accepted to increase the rate of glucose uptake by activating AMPK in skeletal muscles [26]. These data suggested that EFA may employ different signaling pathway from OA to promote glucose uptake into the cells. We therefore examined whether AMPK signaling pathway is involved in glucose uptake induced by EFA and found that EFA markedly induced phosphorylation of AMPK and its downstream ACC. In contrast to EFA, OA had no significant effect on the activation of AMPK and ACC, which is consistent with findings reported previously [27]. We showed that EFA increased AMPK and ACC in C2C12 cells, to our knowledge, which has not been reported previously. We further confirmed that the stimulatory effect of EFA on glucose uptake through the AMPK activation by examination in the presence of araA. A competitive inhibitor of AMPK, araA has been widely used to assess the roles of an AMPK pathway in various cellular processes [28,29]. We found that araA inhibited AMPK/ACC phosphorylation and significantly reduced EFA-stimulated glucose uptake, indicating that AMPK is definitely involved in the stimulatory effect of EFA on glucose uptake in C2C12 cells. Several environmental and nutritional stresses that increase the intracellular AMP/ATP ratio cause AMPK activation [11,12]. In addition, AMPK can also be activated by natural polyphenols, curcumin, many natural products, and some conjugated linoleic acids (CLAs) [22,30,31]. Especially, CLAs, isomers of LA naturally present in ruminant dairy products, have been reported to attenuate PA-induced insulin resistance through the activation of AMPK, PI3K, and AS160 in C2C12 myotubes [17,18]. On the other hand, AMPK increases fatty acid oxidation through the sequential activation of ACC. Accordingly, EFA-induced ACC phosphorylation may contribute to ameliorating the PA-induced impairment of glucose uptake in part through lowering the intracellular accumulation of deleterious lipid metabolites such as diacylglycerol by fatty acid oxidation. Further studies should be required to elucidate the precise mechanism underlying the stimulating effect of EFA on glucose uptake in skeletal muscle cells and its biological significance.
In conclusion, we showed in this study that EFA plays a stimulatory role in glucose uptake in differentiated C2C12 skeletal muscle cells. This beneficial effect is associated with the ability of EFA to restore a suppressed insulin signaling pathway at PA-induced insulin resistant condition and stimulate AMPK pathway in C2C12 cells.
Diabetes mellitus (DM) is one of the most common metabolic syndromes in which a patient has high blood glucose. For the most part of DM is type 2 DM, which is mainly characterized by insulin resistance. For the effective treatment of type 2 DM, nutritional regulation and exercise are usually recommended in an attempt to increase insulin sensitivity. In this regard, the observations in this study may have potential nutritional implications. Increasing EFA intake may be good nutritional management to improve insulin responsiveness, and therefore improve insulin resistance.
Go to :
 XML Download
XML Download