

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
A good experimental animal model is vital to understand the underlying mechanisms of the particular disease [1,2]. A suitable experimental animal model is also an ideal platform for the testing of newly developed drugs prior to human use for the possible cure of the target disease [2,3].
The neurotoxic effect of 1-methyl-4-phenyl-1,2,3,6-tetra hydropyridine (MPTP) on dopaminergic neurons at substantia nigra has been established in various animals [3,4]. MPTP has been used to induce symptoms of Parkinson's disease (PD) in a number of animals. Repeated MPTP administration over several days produces a Parkinsonian syndrome, such as decreased excise abilities, neurochemical changes, and histopathological differences [5].
The active accumulation of 1-methyl-4-phenylpyridinium (MPP+) in dopaminergic terminals via the dopamine transporter occurs because of the specific neurotoxic action of MPTP on dopaminergic neurons [6,7]. The oxidation of MPTP to MPP+ by monoamie oxidase B (MAO-B) is essential to the neurotoxic effect, especially at the substantia nigra. It has been suggested that the toxic effects of MPP+ are related to its accumulation within mitochondria [8], its inhibition of mitochondrial respiration at complex I [9,10], and the depletion of ATP in the affected cells [11]. The endogenous levels of dopamine, as a biogenic amine, are involved in emotion, reward systems, and motor control in human [12,13]. Tetrahydrobiopterin is an essential cofactor of tyrosine hydroxylase, a rate limiting enzyme for synthesis for dopamine synthesis [14,15].
Differences in susceptibility to MPTP by different animal species have been recognized [6,7,16]. Mice are sensitive to the neurotoxic effects of MPTP, with primates being more sensitive [17,18]. Although several mouse strains have been used for the development of a Parkinsonian animal model by the repeated injection of MPTP [6,7,16,19], the differential susceptibilities between mouse strains following MPTP injection as measured by behavior changes and alteration in the content of the endogenous dopamine, tetrahydrobiopterin, and tyrosine hydroxylase have not been addressed.
In this study, we evaluated three mouse strains in MPTP-induced PD by determining the endogenous concentration of tetrahydrobiopterin and dopamine, and the expression levels of tyrosine hydroxylase in brain regions. We directly quantified the endogenous levels of dopamine and tetrahydrobiopterin at the striatum three commonly-used mice strains - C57BL/6, Balb-C, and ICR - using liquid chromatography-tandem mass spectrometry (LC-MS/MS) after MPTP administration [20]. We also measured the striatum expression levels of tyrosine hydroxylase in the mouse strains and evaluated behavioral changes after subcutaneous (s.c.) MPTP administration using the Rota-rod test and an open field behavior recording system. The results revealed differences in behavior and dopamine level in the brains of the mouse strains.
Go to : 
METHODS
Materials
(6R)-5,6,7,8-Tetrahydrobiopterin dihydrochloride (BH4 2HCl), dopamine hydrochloride (dopamine HCl), MPTP, monoclonal anti-tyrosine hydroxylase antibody, and monoclonal anti-β-actin antibody were purchased from Sigma-Aldrich (St. Louis, MO, USA). Epilson-acetamidocaproic acid (AACA) was donated by Kuhnil Pharmaceuticals (Seoul, Korea). T-PER® tissue protein extraction reagent and protease inhibitor cocktail were purchased from Thermo Scientific (St. Louis, MO, USA). IRDye® 800CW conjugated goat (polyclonal) anti-mouse IgG was purchased from LI-COR® Biosciences (Lincoln, NB, USA).
Animal treatment and sample preparation
All procedures in this study were performed according to protocol approved by the Institutional Animal Care and Use Committee at Dankook University. Six-week-old C57BL/6, Balb-C, and ICR mice were purchased from Daehan-Bio Link (Seoul, Korea). They were housed in cages in groups of 10 or 11 animals under controlled environmental conditions (23±2℃; relative humidity 50±10%; 12:12-h light-dark cycle until the initiation of the experiment; food and water ad libitum) before use in experiments. Each mouse was injected subcutaneously (s.c) with MPTP (20~30mg/kg). After testing by Rota-rod and open field test systems as detailed below, mice were killed by cervical dislocation, and each brain was quickly retrieved. The striatum, substantia nigra, and hippocampus were quickly dissected on ice under a dissecting microscope. The dissected brain regions were quick-frozen in a deep-freezer (-80℃) until further analysis.
LC-MS/MS
Preparation of stock solution, calibration standards, quality control, and sample preparation for LC-MS/MS system were done as previously described [20]. The liquid chromatography system was an Accela system (Thermo Fisher Scientific, Waltham, MA, USA) equipped with a nanospace SI-2 3133 solvent delivery module as an auto-sampler (Shiseido, Tokyo, Japan), and connected to a Discovery Max (Thermo Fisher Scientific) quadrupole tandem mass spectrometer coupled with electrospray ionization (ESI-MS/MS). Chromatographic separation was achieved by using hydrophilic interaction chromatography (HILIC) using a Sepax Polar-Imidazole (2.1×100 mm internal diameter, 3 µm particle size) high-pressure liquid chromatography (HPLC) column (Sepax Technologies, Newark, DE, USA) with a 4×2 mm C18 guard column (Phenomenex, Torrance, CA, USA). The ESI mass spectrometer was operated in the positive ion mode. Multiple reaction monitoring (MRM) of the precursor-product ion transitions was m/z 242.1 → m/z 166.0 for BH4, m/z 154.1 → m/z 90.0 for dopamine, and m/z 174.1 → m/z 114.0 for AACA. The optimized ESI conditions could sensitively detect BH4, dopamine, and IS with positive ion detection mode. The most abundant protonated ion peaks in the Q1 mass spectra of BH4 and dopamine were at 242.1 m/z and 154.1 m/z, respectively. There was no evidence of fragmentation and adduct formation. The product ions ([M+H]+) in the Q3 mass spectra and proposed fragmentation patterns were BH4 at m/z 166.0 (2-amino-7,8-dihydropteridin-4(1H)-one) by loss of propane-1,2-diol, and dopamine at m/z 90.9 (butane-1,2-diol) by loss of (E)-3-methylpent-3-en-1-amine. The interpretation and analysis of data followed previously published methods [20].
Rota-rod test
Mice were trained for two consecutive days to stay on a rota-rod (Ugo Basile, Comerio VA, Italy) at least for 600 seconds (16 rpm, fixed speed) before the start of testing. On the test day, baseline data was collected and then mice were evaluated for their latency to fall after s.c administration of 20~30 mg/kg of MPTP. Rota-rod testing was performed twice per mouse with an intervening interval of at least 10 minutes. The times in seconds from the two tests were averaged.
Open field behavior test
Each mouse was put in a white or black plastic rectangular box (40×27×27 cm) after s.c administration of 20~30 mg/kg MPTP. The mice injected with the same volume of saline were used as the control group. The mice were observed with a CCD camera connected to a recording system. The recorded behavior was analyzed by EthoVision Version 2.3 (Noldus information Technology, Wageningen, The Netherlands). An automated video tracking system was used for calculation of moving distances and rearing frequencies. The mice were observed for 6 hours in normal lighting. The white or black plastic rectangular box was cleaned with 70% alcohol and water between trials.
Western blotting
Quick-frozen tissue samples were homogenized and analyzed via immunoblotting as previously described [21]. Briefly, the homogenized samples were boiled in Laemmli sample buffer and the proteins separated by 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis. The resolved proteins were transferred to a polyvinylidene difluoride membrane (Millipore, Billerica, MA, USA). Protein transfer was confirmed by staining membranes with Ponceau Red. Each membrane was incubated with an appropriate primary antibody for overnight at 4℃. After extensive washing with Tris-buffered saline containing Tween 20, each membrane was incubated with the appropriate secondary antibody. Densitometry of protein bands was performed with the software for Odyssey Infrared Imaging System (LI-COR Biosciences).
Data analysis
All values given in the text are mean±SEM. Differences between means were evaluated using a Student's t-test. For comparison between strains, Two-way ANOVA with the factors "strain" (ICR, Balb C, and C57BL/6) and "day after treatment" (1, 2, and 3 days) were computed using Prism (version 5.0) (GraphPad Software, San Diego, CA, USA). Post-hoc analysis of significant differences was performed using Bonferroni post-tests for multiple comparision. Statistical significant differences were taken at the p<0.05 level. The n values given represent numbers of tissues or animals used in each experiment.
Go to :
RESULTS
Rota-rod performance is decreased but recovers after MPTP administration
C57BL/6 mice were trained to stay on the Rota-rod for 600 seconds when the system was operating at a fixed rate of 16 rpm. The trained animals were injected with MPTP (30 mg/kg s.c.) and then evaluated their latency to fall off the Rota-rod. Fall latency 6 hours after injection was significantly decreased (Fig. 1A). The decreased latency persisted until 24 hours post-injections, although the decrease was significantly greater at 6 hours than at 18 hours (Fig. 1A). Similar findings were obtained with ICR mice (Fig. 1B). However, the latency to fall of ICR mice was greater than for C57BL/6 mice 24 hours after injection.
 | Fig. 1Effects of MPTP (30 mg/kg, s.c) on Rota-rod performance. Animals were trained to stay on an operating Rota-rod for 600 seconds. On the test day, animals were evaluated on the Rota-rod 6 and 24 hours after MPTP administration. (A) MPTP (C57BL/6, 30 mg/kg, s.c.), (B) MPTP (ICR, 30 mg/kg, s.c.), (C) MPTP (C57BL/6, ICR, Balb C, 20 mg/kg, s.c.), (D) body weight after MPTP administration. Data are expressed as mean latency to fall off the Rota-rod (vertical bars represent standard error of the mean). *p<0.05, **p<0.01 vs 0 hour (naïve) group, #p<0.05 compared between 6 hours and 24 hours groups by Student's t-test, n=6~10.
|
Multiple injections of MPTP (30 mg/kg, s.c.) were given at 24 hour intervals and mice were tested their behavioral changes (Fig. 2). This treatment produced extensive mortality and the experiment was terminated day 2, so the MPTP dosing regimens have to be changed. To limit mortality, the three different mouse strains were treated with a lower dose of MPTP (20 mg/kg, s.c), and with multiple injections of this lower dose, and were evaluated for their latency to fall on the Rota-rod. However, the injection of lower dose of MPTP (20 mg/kg, s.c.) didn't decrease the latency on rota-rod at 24 hours after MPTP injection (Fig. 1C). Moreover, multiple injection of MPTP (20 mg/kg, s.c.) at 24 hours intervals for 3 days didn't significantly decrease latency to fall on a rota-rod (data not shown). There were no significant changes in the body weights of the examined mice during the 3-day period of MPTP injection and 24 hours after MPTP injection in all three mouse strains (Fig. 1D).
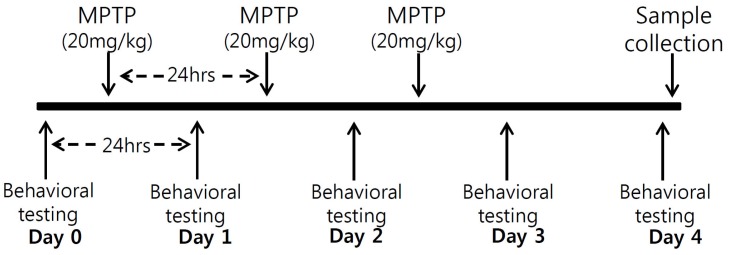
MPTP injections decrease moving distances and rearing frequencies
We further evaluated the mice after MPTP injections with alternative methods to see whether the lower dose of MPTP was insufficient to be toxic to dopaminergic neurons or whether the Rota-rod test was not sensitive enough to detect behavioral changes. To assess the behavioral effects, the movements of C57BL/6, Balb C, and ICR mice after injecting MPTP (20 mg/kg, s.c.) were evaluated by the open field test for 6 hours. The total moving distances recorded and analyzed until 1 hour after MPTP injection were significantly decreased at day 2 and day 3 in C57BL/6 mice (Fig. 3A). However, the moving distances were still recovered after 24 hours (day 4) similarly as day 0 if the mice were not treated with MPTP (Fig. 3A).
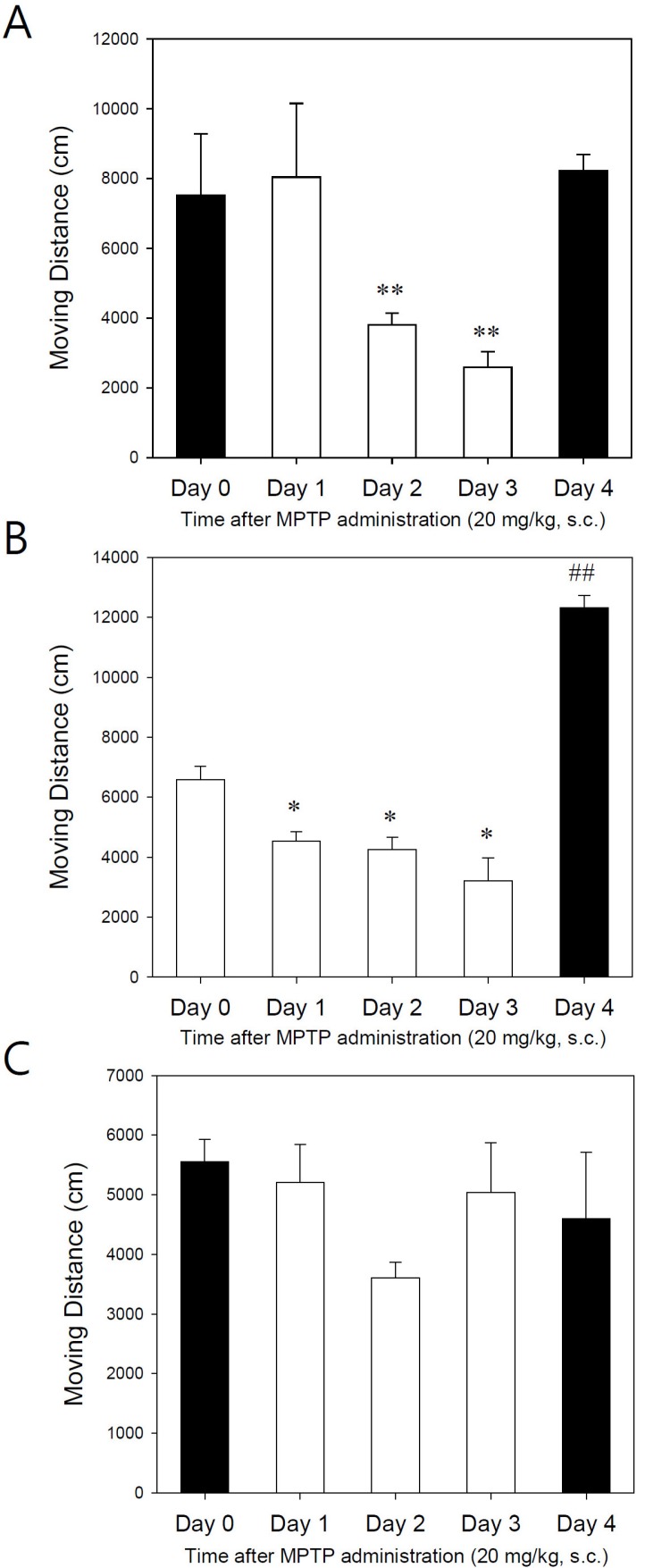 | Fig. 3Effect of MPTP (20 mg/kg, s.c) on moving distance for 1 hour following MPTP administration. (A) C57BL/6 mice, (B) Balb C mice, (C) ICR mice. Data are expressed as accumulated total moving distances in cages (vertical bars represent standard error of the mean). *p<0.05, **p<0.01, ##p<0.01 vs day 0 (n=6~10).0
|
In parallel, Balb C mice were similarly tested. The total moving distances were significantly decreased from day 1 and further decreased following the repeated injection of MPTP (20 mg/kg, s.c.). Moreover, the total moving distances at day 4 (without MPTP injection) were markedly increased by two-fold compared with day 0 (Fig. 3B). The observations were consistent with the hyperactivity of the mice at day 4. Of interest, ICR mice did not respond well at the lower dose of MPTP (20 mg/kg, s.c.) in the same experiments and showed no specific changes at day 4 (Fig. 3C).
Concerning rearing frequencies, all three strains of mice responded well to the MPTP injection (Fig. 4). The rearing frequencies until 1 hour after MPTP injection were significantly decreased from day 1 and its effects lasted until day 3. But, this decrease observed with multiple MPTP injections recovered after 24 hours (day 4) with a frequency similar to that noted on day 0 if the mice were not injected with MPTP (Fig. 4).
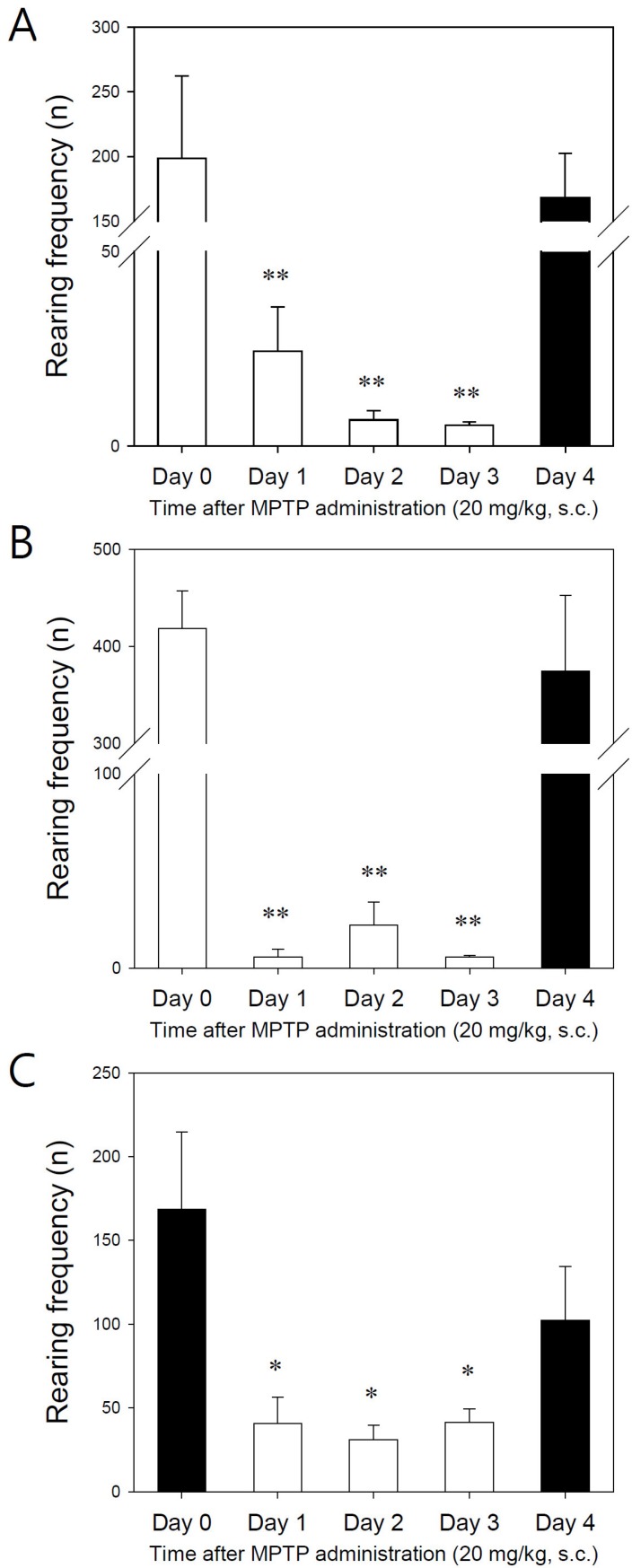 | Fig. 4Effect of MPTP (20 mg/kg, s.c) on total rearing frequencies until 1 hour after MPTP administration. (A) C57BL/6 mice, (B) Balb C mice, (C) ICR mice. Data are expressed as accumulated number of rearing frequencies in cages (vertical bars represent standard error of the mean). *p<0.05, **p<0.01 vs day 0 (n=6~10).
|
We further analyzed the data until 3 hours and 6 hours in the same experiments. The results showed a very similar pattern at both times in the total moving distances (Table 1) and rearing frequencies (Table 2). The total moving distances were significantly decreased following the repeated injection of MPTP (20 mg/kg, s.c.) in both C57BL/6 and Balb C mice. At the same time, the total moving distances at day 4 (without MPTP injection) were recovered to similar distances [22]. Similarly, ICR mice did not display significant differences at 3 hours or 6 hours evaluation when the lower dose of MPTP was used. Interestingly, Balb C mice at day 4 moved an extended distance (up to 3-fold) compared to day 0. These data indicated that mice, especially Balb C mice, are hyperactive during the recovery time at day 4 (Table 1).
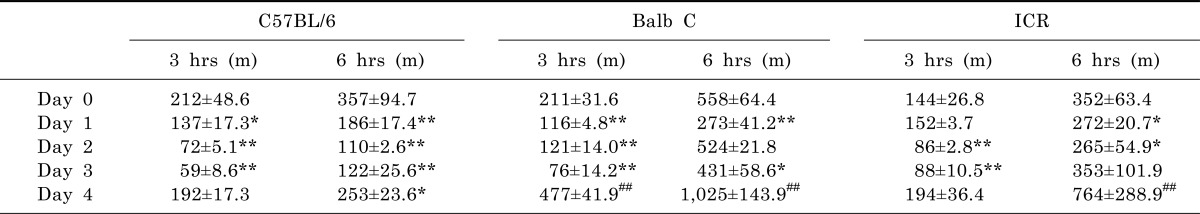
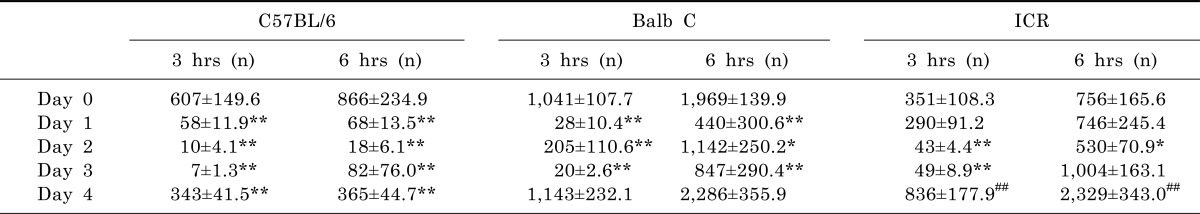
All three strains of mice displayed significantly decreased rearing frequencies from day 1 to day 3, but still showed robust recovery of activities at day 4 (Table 2). Especially, the rearing frequencies at day 4 were markedly increased up to 5-fold in ICR mice. But, there were no increased rearing activities at day 4 either in C57BL/6 or Balb C mice. These data suggest that MPTP can differentially affect the rearing behavior of different mouse strains.
Although, repeated administrations of MPTP significantly affected the moving distances and rearing frequencies in all three mouse strains, we further analyzed the data to see mice strains are differently susceptible (Fig. 5). In both parameters, ICR strain is statistically less sensitive to MPTP than C57BL/6 or Balb C strains.
 | Fig. 5Comparison of moving distances and total rearing frequencies among three mouse strains Effect of MPTP (20 mg/kg, s.c) on moving distances (A) and total rearing frequencies (B) until 1 hour after MPTP administration over three days. In each case, the mean results were normalized by the mean of day 0. Vertical bars represent standard error of the mean and negative SE bars have been deleted for clarity. Two-way ANOVA, *p<0.05, **p<0.01 vs ICR (n=6~10).
|
MPTP injections decrease endogenous dopamine levels, but not tetrahydrobiopterin, in mouse brain
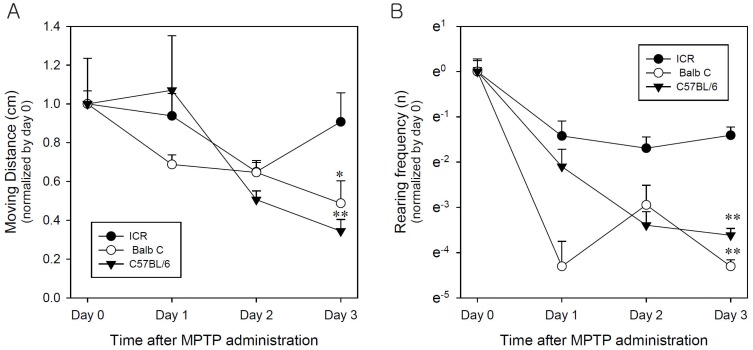
MPTP destroys dopaminergic neurons in the substantia nigra, which causes Parkinsonism [11,16]. Recently, we reported a direct detection method for tetrahydrobiopterin (BH4) and dopamine in rat brain using LC-MS/MS [20]. Dopamine and BH4 were presently measured in brain regions of the three strains of mice after administration of MPTP (Fig. 6 and 7). Three injections of MPTP (20 mg/kg s.c.) for a 24 hours interval resulted in an approximately 60% decrement of dopamine in the striatum in all three strains (Fig. 6A). In parallel, endogenous levels of dopamine in the three strains of mice were measured in the substantia nigra (Fig. 6B) and in the hippocampus (Fig. 6C). The three injections of MPTP did not statistically affect the substantia nigra in any of the strains (Fig. 6B). But, the dopamine concentration in the hippocampus of C57BL/6 mice was slightly decreased by the multiple MPTP injections (Fig. 6C).
 | Fig. 6Effects of MPTP (20 mg/kg, s.c) on dopamine concentration in mouse brain section. The endogenous level of dopamine in mice brain scetions ((A) striatum, (B) substantia nigra, (C) hippocampus) were measured by LC-MS/MS in three different mice species. (D) internal calibration for dopamine. Data is expressed as mean±s.e.m. *p<0.05, **p<0.01 (n=5).
|
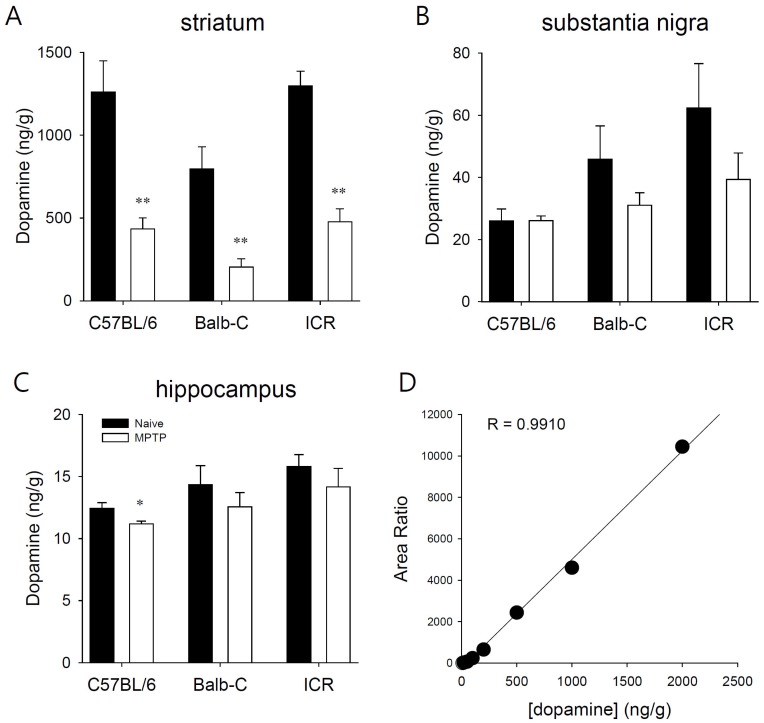
 | Fig. 7Effects of MPTP (20 mg/kg, s.c) on tetrahydrobiopterin (BH4) concentration in brain regions. The endogenous level of BH4 in mice brain regions ((A) striatum, (B) substantia nigra, (C) hippocampus) were measured by LC-MS/MS in three different mice strains. (D) Internal calibration for BH4. Data is expressed as mean±standard error of the mean (n=5).
|
BH4 is an essential cofactor for dopamine synthesis through its action on dopamine hydroxylase. BH4 has been intensively studied concerning the development and progression of PD [23-26]. The endogenous levels of BH4 in brain section were measured after three doses of MPTP (20 mg/kg s.c.) delivered at 24 hours intervals. No statistically significant differences were evident in the striatum, substantia nigra, and hippocampus among the three strains of mice (Fig. 7). These data suggested that the administration of MPTP can decrease the endogenous level of dopamine in mouse brain by destroying dopaminergic neurons, mainly at striatum, with no effect on BH4 levels in mouse brain.
MPTP injections decrease expression of tyrosine hydroxylase in mouse striatum
Based on the assay of dopamine and tetrahydrobiopterin at striatum, substantia nigra, and hippocampus by LC-MS/MS, the dopamine concentration was greatest in the striatum. To ascertain the activity of tyrosine hydroxylase following MPTP administration, we quantified the total expression levels of tyrosine hydroxylase at striatum in the three strains of mice (Fig. 8). The expression levels of tyrosine hydroxylase in the striatum were significantly decreased by the systemic administration of MPTP (20 mg/kg, s.c.) in all three strains. However, the tyrosine hydroxylase decrease was least for ICR mice.
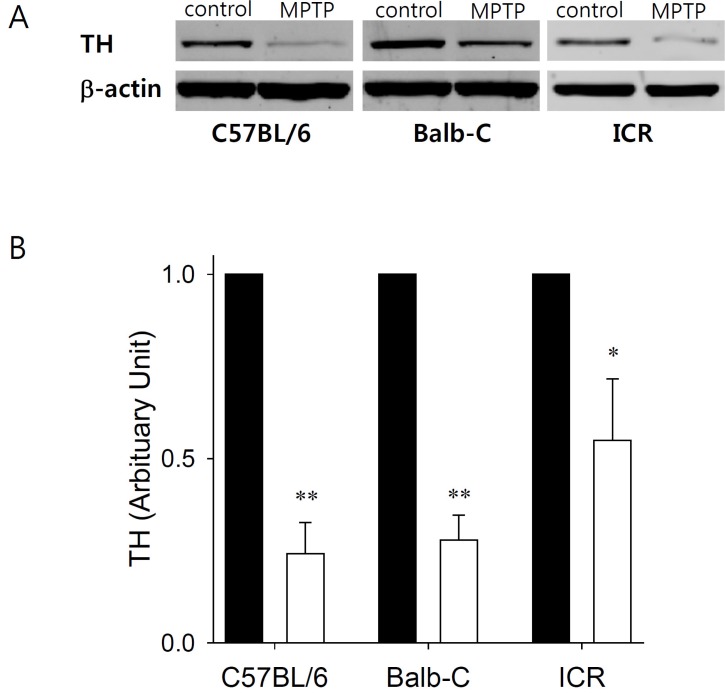 | Fig. 8Effects of MPTP (20 mg/kg, s.c) on the expression of tyrosine hydroxylase in the striatum in the three strains of mice. (A) Representative immunoblots of tyrosine hydroxylase (TH) and β-actin expression. (B) Densitometry plot of TH expression immunoblots normalized to the control levels, with and without MPTP treatment (n=4). *p<0.05 and **p<0.01 vs. black bar (control group).
|
Go to :
DISCUSSION
Development of an animal model for the specific disease under investigation is very important to further the understanding of the underlying mechanism of disease, and provides the chance to test newly developed new drugs before their human application [1,2]. However, the plethora of existing animal models can make the decision of the most appropriate model challenging. It is true for investigating the interactions in brain on the pharmacological and/or toxicological effects of certain agents that link to delineate neural mechanisms underlying in a specific disease [27].
PD is a progressive neurodegenerative disease caused by the destruction of dopaminergic neurons in the substantia nigra [28]. The underlying mechanisms are still not fully understood [5]. Moreover, the current approach to the treatment of PD involves suppressing disease progression rather than achieving a cure [28]. This approach is unsatisfactory, given the high prevalence of PD worldwide, with its concomitant morbidity and mortality; improved understanding of the underlying mechanisms and novel therapeutics are crucial [2,3].
Neuromelanin-containing dopamine neurons in the substantia nigra projecting to the striatum in the brain are selectively degenerated in this disease [29]. In PD animal model, several agents effective against dopaminergic neurons are available [30]. Of these, the administration of MPTP is an established and valid method to induce PD symptoms in mice [3,4]. The effect of the parkinsonism-inducing neurotoxin 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine on central catecholamine neuron in C57BL/6 mice revealed that MPTP caused a severe reduction of endogenous DA in substantia nigra and striatum which was following by an increase in the 3,4-dihydroxyphenylacetic acid (DOPAC)/dopamine ratio [27]. But, little is known about MPTP susceptibilities between different mice strains. C57BL/6, Balb-C, and ICR strains of mice, among others, have been used to develop disease models. However, there is limited information about the different responses among those mice strains after injection of MPTP, which hinders research concerning the underlying mechanisms of PD.
In this study, we evaluated behavioral changes after subcutaneous injection of MPTP using the Rota-rod test and open field behavior recording. In parallel, we evaluate the MPTP-induced mouse model of PD by determining the levels of BH4 and dopamine in brain regions of three selected mouse strains. BH4 is an essential cofactor for hydroxylation of cyclic amino acids including dopamine catalyzed in a rate limiting fashion by tyrosine hydroxylase [14,15]. Recently, we reported a novel method for direct detection of BH4 and dopamine in rat brain using LC-ESI-MS [20]. Here, the method was applied to the MPTP-induced mouse models of PD to monitor the changes of dopamine concentration and the levels of endogenous BH4. We directly quantified the endogenous levels of dopamine in ICR, C57BL/6, and Balb-C mouse strains using LC-MS/MS. The changes of neurotransmitter concentration in several brain regions are influential in the development of a variety of psychiatric and neurodegenerative diseases [25,31]. Therefore, it is clearly necessary to precisely determine the level of dopamine as a means of diagnostically evaluating PD and for the screening of potential therapeutic products to modulate dopamine levels [32].
Presently, repeated administration of MPTP affected exercise abilities of mice, including moving distances and rearing frequencies. However, the generally used methods for evaluation of motor-activities appeared to be too insensitive to detect the fine behavioral changes in the Parkinsonian mice models. The endogenous dopamine concentrations were significantly decreased after repeated MPTP injection, but BH4 was unchanged in mouse brain regions. In addition, the expression levels of tyrosine hydroxylase in the striatum were significantly decreased by MPTP injection. These results suggest that the decreased levels of dopamine in the striatum are mainly due to the extensive damage of dopaminergic neurons in this region. Consistent with previous reports [3,6,7,33], marked differences were evident in the sensitivity of the three strains of mice to MPTP, although the reasons for the strain-related differences in response to MPTP remain unclear. The fundamental difference in the sensitivity to toxic MPTP may be one possibility.
In conclusion, we tested the effects of repeated MPTP injedctions, an animal model of Parkinson's disease, in three strains of mice on rota-rod performance, locomotor activity, as well as striatal, hippocampal, and substantia nigra levels of dopamine and tetrahydrobiopterin and striatal levels of tyrosine hydroxylase. Our results showed that the ICR strain was generally less sensitive to MPTP on rota-rod performance, locomotor suppression, and striatal tyrosine hydroxylase suppression. We found that C57BL/6 and Balb-C mice were more sensitive to the dopaminergic neuronal toxicity of MPTP than was ICR mice based on open field test, tyrosine hydroylase expression.
Go to :
 XML Download
XML Download