

 PDF
PDF ePub
ePub Citation
Citation Print
Print


ABBREVIATIONS
Akt
Protein Kinase B (PKB)
AMPK
5' adenosine monophosphate (AMP)-activated protein kinase
ASIC
acid-sensing ion channel
ENaC (ENaC-α)
(α-subunit of) epithelial Na+ channel
ERK
extracellular-signal-regulated kinases or classical MAP kinases
HS
hypoxic solution
KO2 channels
oxygen-sensitive K+ channels
NMDG
N-methyl-D-glucamine
N*oP
channel activity
PI3K
phosphatidylinositol 3-kinase
PI(s)
phosphoinositide(s)
PI(3,4)P2
phosphoinositol 3,4-biphosphate
PI(3,4,5)P3
phosphoinositol 3,4,5-triphosphate
Po
open probability
pS
picoSiemen (10-12 Siemen)
TTX
tetrodotoxin
INTRODUCTION
Hypoxia induces critical stress to cells under ischemic conditions, resulting in membrane depolarization, increased cellular Ca2+, alterations in pH, and finally cell death without recovery to normoxia [1-3].
Upon exposure to hypoxia, chemo-sensitive cells, such as those in the carotid body and adrenal medulla, rapidly elicit appropriate changes in both signal and cell function [4]. Oxygen-sensitive K+ (KO2) channels have been identified in rat pheochromocytoma (PC12) cells from the adrenal medulla [5,6]. Inhibition of this current by hypoxia results in cellular depolarization as well as increased intracellular Ca2+ through voltage-operated Ca2+ channels [4-7]. The Kv1.2 gene was suggested to encode the α subunit of slow-inactivating KO2 channels in PC12 cells [7]. Thus, PC12 cells have been used as a model system to study O2-sensitive channels and their function in response to low O2 levels.
Although KO2 channels are considered to be the main modulator of depolarization in response to hypoxia, other types of ion channels may be involved. For example, hypoxia could potentially increase the background inward Na+ current. Therefore, the depolarization induced by hypoxia may not be solely due to the inhibition of K+ efflux but also the net influx of cations largely carried by Na+ [8]. Thus, although the initial cause of depolarization is reduction of K+ current, TTX-sensitive Na+ influx is critical for depolarization as it is unaffected by hypoxia [9,10].
Recently, amiloride-sensitive and voltage-independent Na+ channels such as ENaC, which currents are enhanced by the extracellular pH drop, have been suggested as the proton sensor involved in the transduction of ischemic signals in CNS neurons [11]. In addition to their pathophysiological role, ENaC-like Na+ channels are tightly regulated in non-epithelial cells in order to avoid cell death due to Na+ loading and a reduced electrochemical membrane gradient [12].
Depolarization through ENaC activated by hypoxia/ischemia remains unclear. A number of signaling pathways that modulate ENaC activity in response to hypoxia have been described, including the Ca2+-calmodulin pathway, AMPK pathway, ERK1/2 pathway, stress-activated protein kinase (also known as the p38 kinase) pathway, and phosphatidylinositol 3-kinase (PI3K)-Akt pathway [13].
Among them, the PI3K/Akt pathway has diverse effects in response to hypoxia. PI3K mediates neuroprotection against cerebral infarction [14], and it produces phosphoinositides, especially PI(3,4)P2 and PI(3,4,5)P3, both of which activate ENaC [15]. As an endogenous marker of tumor hypoxia [16], IκB kinase-β directly interacts and thereby augments ENaC activity [17]. These prior studies suggest that ENaC mediates Na+ influx for the maintenance or induction of depolarization in response to hypoxia. Unfortunately, there has been no report on neuronal ENaC activated by onset of hypoxia.
In this study, we observed that acute hypoxia activated ENaC, which depolarized cells by direct influx of Na+, in rat PC12 cells, as well as reduced K+ outflow through an oxygen-sensing K+ channel.
METHODS
Cell culture
PC12 cells were cultured in RPMI-1640 (GIBCO) medium containing 10% fetal bovine serum, 5% horse serum, 100 U/ml of penicillin, and 100 µg/ml of streptomycin under 5% CO2 at 37℃, and they were transferred every 2~3 days. For electrophysiological recording, cells were subcultured in a 35-mm Petri dish containing round coverslips with a 10 mm diameter.
Solutions
For electrophysiological measurements, standard bath and pipette solutions contained 120 mM NaCl, 5 mM KCl, 1 mM MgCl2, 1.8 mM CaCl2, 25 mM NaHCO3, 1.2 mM KH2PO4, and 10 mM HEPES, adjusted to pH 7.4 using NaOH or HCl. To decrease Na+ content in the pipette solution, NaCl was substituted using 120 mM NMDG-Cl. To test ion selectivity, bath solution contained 100 mM potassium glutamate, 8 mM KCl, 5 mM NaCl, and 10 mM HEPES.
Hypoxic conditions
Hypoxic conditions were generated by bubbling bathing solution with mixed gas containing 95% N2 and 5% CO2. The oxygen concentration was measured with an O2-sensing microelectrode and analyzed as mmHg unit and percentage.
Calcium imaging
For calcium imaging, 2 day-cultured cells were loaded with 5 µM Fluo-3 AM (Invitrogen) and incubated for 30 min at 37℃ in bath solution containing 120 mM NaCl, 5 mM KCl, 1 mM MgCl2, 1.8 mM CaCl2, 25 mM NaHCO3, 1.2 mM KH2PO4, and 10 mM HEPES (pH 7.4). Intracellular Ca2+ was analyzed by the Fluo-3 AM fluorescence dye detection method using commercial software (MetaMorph V7.7, Molecular Devices, USA).
Electrophysiology
Single-channel currents were recorded using a patch-clamp amplifier (Axopatch 200B, Molecular Devices, USA). Patch pipettes were made from borosilicate glass capillaries (Warner, USA) coated with Sylgard elastomer (Dow Corning, USA). Single-channel currents were sampled at 5 kHz and filtered at 2 kHz using an analog-digital interface (Digidata 1440A, Molecular Devices, USA). Voltage pulse was modified from the ramp protocol used by Choi et al. [18]. Experiments were performed at room temperature. Currents were analyzed using Clampfit software (V10.3, Molecular Devices, USA).
RT-PCR
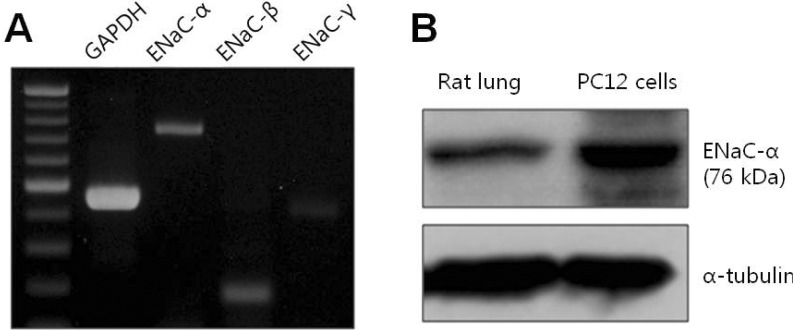
Total RNA was isolated from PC12 cells using TRIZOL reagent (Invitrogen) according to the manufacturer's protocol. One microgram of total RNA was reverse-transcribed using SuperScript II RT (Invitrogen), 1µl of oligo dT primer, and 2.5 mM dNTPs at 50℃ for 1 h. cDNA was amplified using the following primer sequences: rENaC-α, 5'-AATCCTTACAGATACACTGAA-3' and 5'-TTCCTTTCTCATACTGATGGA-3'; rENaC-β, 5'-CAAGAAGAAGGCCATGTGGT-3' and 5' GTACTGGAAGGGGCTGGAAT-3'; rENaC-γ, 5'-CAAAGATCCATCATGGAGAGC-3' and 5'-ATTGGTCAACTGAGTGTCTGT-3'.
Protein extraction and Western blotting
Lung tissue from rat was lysed with RIPA buffer (50 mM Tris-HCl, pH 7.4, 150 mM NaCl, 5 mM EDTA, 1 mM PMSF, and 1% NP-40) containing protease-inhibitor cocktail and homogenized three times for 5 min in a homogenizer (FastPrep-24). PC12 cells were lysed with the same buffer. Whole cell lysates were incubated on ice for 30 min and cleared at 20,000×g for 20 min at 4℃. Proteins were separated by 8% SDS-PAGE and blotted onto PVDF membranes. Blots were incubated overnight at 4℃ using ENaC-α antibody (1:250; Santa Cruz Biotechnology).
RESULTS
To confirm the hypoxic conditions induced by the hypoxic solution (HS) saturated with 95% N2 and 5% CO2, changes in intracellular Ca2+([Ca2+]i) and oxygen levels in PC12 cells were compared with those induced by CoCl2, which is widely used as a chemical hypoxic agent.
When 200 µM CoCl2 was applied for 3 min, [Ca2+]i was significantly elevated (266.4±39.2% compared to control, n=5). Similarly, N2 mixed gas increased [Ca2+]i about 2.8 fold within 3 min (280.2±10.0%, n=6) following bubbling (Supplementary Fig. 1). The oxygen level was remarkably reduced to about 40% within 3 min following onset of HS perfusion (normal: 152.76 mmHg and hypoxia: 91 mmHg, n=3). However, N2 gas did not alter the pH level in the experimental solution. These results show that application of N2 gas induced acute hypoxia.
Acute hypoxia-induced channel with 30-pS conductance
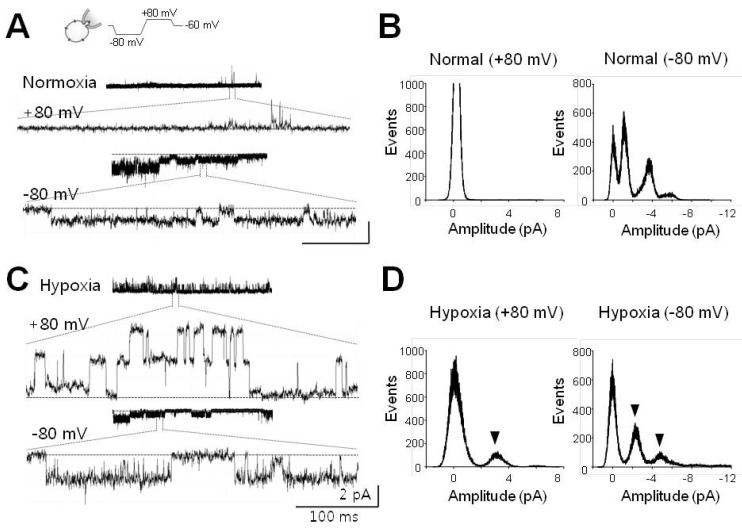
In this study, we tried to identify a channel responsible for direct depolarization in response to acute hypoxia. For activation of such channels, single channel currents were recorded before and after hypoxic stress using a cell-attached (CA) configuration, which can protect the environment of the channel.
After forming CA configuration under symmetrical Na+-rich (120 mM NaCl) conditions, diverse channels with amplitudes ranging from less than 0.5 pA to ~3.5 pA were observed. Channels could be detected at -80 mV, but not at the depolarization potential (+80 mV, refer to Fig. 1A). Hypoxic stress did not affect channel activities, except for one channel with a conductance of 30-pS and an amplitude of 2.5 pA at +80 or -80 mV (Fig. 1C). This channel was activated ~2 min after application of HS to the bath solution (arrowheads in Fig. 1D). Further, the channel could be detected at a rate of ~13% (one of seven to eight cells tested) under hypoxic conditions, and it showed long-lived opening behavior (mean open time, to 310.7±52.4 ms, n=4) and open probability (Po) was 0.88±0.02 (n=4).
The 30-pS channel has Na+ selectivity
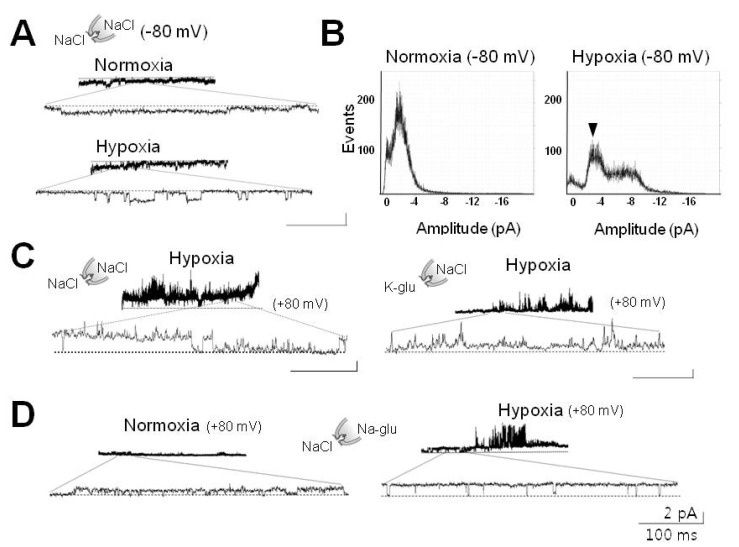
Interestingly, the 30-pS channel was activated in response to acute hypoxia. Patch mode was switched to inside-out (IO) configuration from CA in order to identify which ion passes through. Under symmetrical Na+-rich conditions, channel activation induced by hypoxia was maintained after excision of the membrane patch, implying that the channel was permeable to Na+ or Cl- (Fig. 2A and 2B). This activation disappeared after return to normoxia (data not shown), suggesting that the channel could work in the absence of cytosolic components. In the present study, we did not observe activation of the channel in IO patch mode from CA mode upon exposure to HS.
In the next step, to determine whether or not the channel is permeable to either Na+ or Cl-, the effects of chloride on channel currents were examined using glutamate. Hypoxia-induced 30-pS channels were not affected by removal of Cl- (Fig. 2D). For the experiments shown in Fig. 2C and D, we clamped the cell interior at +80 mV in order to record currents elicited either by internal (bath) K+ or external (pipette) Cl-.
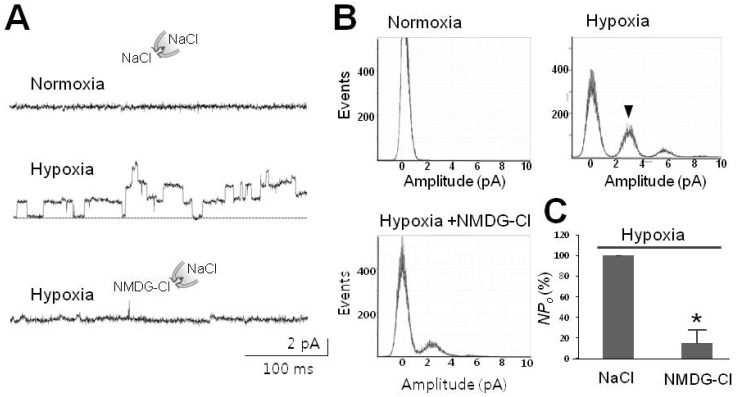
To examine cation selectivity, IO membrane patches were excised from cells containing 30-pS channels activated by hypoxia. Channel activity was switched off by recovery to normoxia (upper trace in Fig. 3A) but vigorously reactivated by subsequent application of HS (middle in Fig. 3A and right panel in 3B). When Na+ was removed by replacement with eqimolar K+ or N-methyl-D-glucamine (NMDG) in the bath solution (i.e. cytoplasmic side), channel activity was completely abolished (Fig. 2C and 3A). As shown in the lower trace in Fig. 3A, channel activity (N*Po) in the presence of NMDG, but not Na+, was remarkably reduced to 18.07±6.13% (n=3) compared to that under hypoxic Na+-rich conditions. Channel inhibition was also observed upon the removal of Na+ in the pipette solution (extracellular side). Since the solutions used in the present study contained Ca2+ (1.8 mM), it is unlikely that Ca2+ passed through the 30-pS channel. As shown in Fig 3A, channel activity was significantly (p<0.001) reduced under Na+-free conditions containing Ca2+. These results suggest that the channel was predominantly Na+-selective.
Finally, to determine the channel preference for Na+, we examined whether or not another monovalent such as K+ passes through. After adjustment to K+-rich (100 mM K-glutamate) conditions, channel activity completely disappeared under low Na+ content (20 mM), as shown in Fig. 2C, suggesting that K+ did not pass through. Therefore, the hypoxia-activated 30-pS channel was predominantly Na+-selective.
The Na+-selective 30-pS channel is blocked by lidocaine and amiloride
Since the channel activated by acute hypoxia in PC12 cells was a Na+ channel, we explored its pharmacology using neuronal Na+ channel blockers such as lidocaine and amiloride. Channel activity was dramatically reduced to 80% by 100 µM lidocaine (Supplementary Fig. 2).
Regarding Na+ channels in neurons, most voltage-independent Na+ channels are rapidly inactivated, with mean open times at best from 16 ms to 0.7 ms [19,20]. On the contrary, voltage-independent Na+ channels such as epithelial Na+ channel (ENaC)/degenerin family members are open longer than voltage-dependent Na+ channels, with relatively high Po up to 0.9 [21]. According to such simple kinetic criteria, we found that the 30-pS channel shared properties with ENaC, including conductance of 30 pS, 310 ms-long mean open time, and Po of 0.88 at -80 mV, as stated with Fig. 1, which are close to those of epithelial α-subunit Na+ (ENaC-α) channel [22].
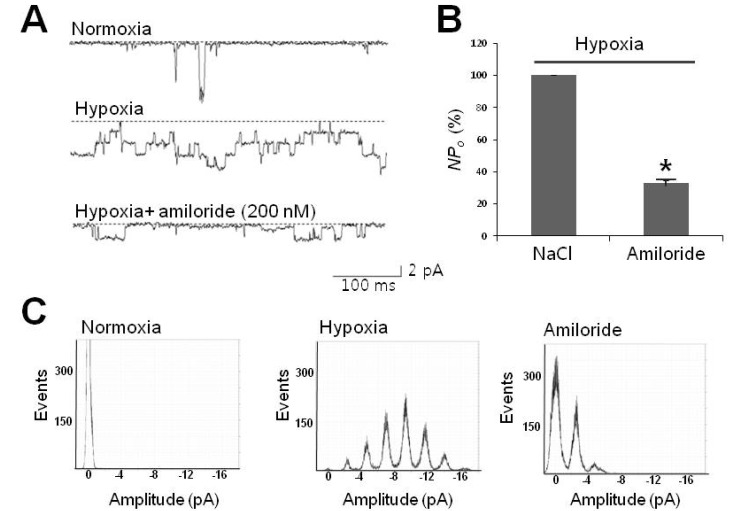
To determine whether or not the 30-pS channel shares common pharmacology with ENaC, we applied amiloride, which is widely used to specifically block ENaC and acid-sensing ion channel (ASIC). As shown in Fig. 4, less than 1 min following treatment with a nanomolar amount of amiloride (200 nM), channel activity was significantly reduced to about 30.58±10.62% (n=3) compared to that of control (p<0.05). Since ASICs require just a few tens of µM amiloride to achieve 50% inhibition (IC50), and also Ca2+-permeability [23], the 30-pS channel was determined to be an ENaC rather than the ASIC family member.
ENaC is endogenously expressed in PC12 cells
Previous results have strongly suggested that the 30-pS channel activated by hypoxia demonstrates properties resembling ENaC. Therefore, ENaC RNA and protein were confirmed by using RT-PCR and Western blotting in order to determine that ENaC is endogenously expressed in PC12 cells.
Using constructed subunit primers of ENaC consisting of α, β, and γ subunits, we confirmed the presence of RNAs corresponding to three subunits in PC12 cells (Fig. 5A). Finally, we demonstrated the endogenous protein expression of ENaC using antibody specific to rat ENaC-α as well as RNA expression using RT-PCR. As shown in Fig. 5B, ENaC protein was detected by Western blotting using ENaC-α-specific antibody and lung tissue as a positive control (left lane in Fig. 5B). These results confirm that the 30-pS channel could be a member of the ENaC family activated by acute hypoxia in rat PC12 cells.
Taken together, the results in the present study suggest the presence of an ENaC-like channel activated by acute hypoxia in neuronal rat PC12 cells.
DISCUSSION
The present study provides evidence for the presence of a Na+ channel activated by acute hypoxia in PC12 cells. This result was supported by the following: the 30-pS chan nel was activated under hypoxic conditions and its activity disappeared upon return to normoxia (Figs. 1 and 2). Second, the channel was selective for Na+, whereas it was unaffected by Ca2+, Cl-, or K+. Finally, the channel was blocked by lidocaine, a neuronal Na+ channel blocker.
In contrast to voltage-gated Na+ channels, which are rapidly inactivated, opening of the hypoxia-activated channels lasted longer than 300 ms, and the inactivation process was not observed when the recording lasted for longer than 10 sec (Po=0.88, refer to Fig. 1C) [21]. The hypoxia-activated channel shared properties similar to those of ENaC, a member of the vertebrate ENaC/ASIC family [24]. On the other hand, this channel revealed differences in kinetics and pharmacology from those of ASIC. Further, it was sensitively blocked with a low dose (200 nM) of amiloride, whereas ASICs are less sensitive (up to hundreds µM) and rapidly inactivated [25,26]. Since the pH level did not respond to bubbling of anoxic N2 gas, it was identified as an ENaC rather than ASIC. This suggestion was supported by the expression of ENaC protein in PC12 cells (Fig. 5), although direct evidence is required that heterologously expressed ENaC is also sensitive to acute hypoxia.
The Na+ channel conductance activated by acute hypoxia was larger than those reported in studies on endogenous ENaCs. The conductance was about 30 pS (2.5 pA at-80 mV) in the present study, whereas a unit conductance of ENaC has been reported at best to be less than 10 pS [27-29]. If the hypoxia-activated Na+ channel belongs to the ENaC family, then a larger conductance would conflict with those of ENaCs which are believed to form a heteromeric channel composed of four subunits α, β, γ, and δ [21]. Only a few reports have reported conductance levels of 21~40 pS from recordings performed in the planar lipid bilayer or on homomeric ENaC-α [30-32], implying that the conductance could be modulated by β- and γ-subunits as well as cytoplasmic factor(s). For instance, reduction of disulfide bonds with dithiothreitol from ENaC incorporated into planar lipid bilayers reduced single channel conductance from 40 pS to 13 pS [30]. Considering this redox modulation of ENaC conductance, it is likely that oxidation can elevate unit ENaC conductance.
The mechanism underlying ENaC activation during acute hypoxia is difficult to explain based solely on this study. At present, there is a general agreement that intermediate or long-term hypoxia inhibits ENaC activity in the alveolar epithelia, whereas the effect of acute hypoxia on neuronal ENaC unclear. This channel is upregulated by proteolytic enzymes and phosphoinositides (PIs), including PI(4,5)P2 and PI(3,4,5)P3. Both PIs can interact with the N terminus of ENaC-β as well as the proximal C terminus of ENaC-γ, thereby enhancing ENaC activity. Depletion of membrane PI(4,5)P2 inhibits ENaC activity [33]. On the other hand, hypoxia provokes intense Ca2+ mobilization as well as the level of PI(4,5)P2, stimulating calpain proteolytic activity [34]. Thus, it can be stated that acute hypoxia activates proteolytic enzymes such as calpain and prostasin, which, in turn, trigger the activation of ENaC [35]. In order to mimic hypoxia, we did not add ATP to the bath. Therefore, the effect of PIs on the channel activity was greatly limited since ATP is essential to the modulation of membrane-delimited PI kinases such as PI3K or PLC. Throughout the experiments, activation of the 30-pS channel was not spontaneous and usually occurred 2~3 min after exposure to hypoxia. These channels in the excised patch were re-activated without any alteration in unit conductance in response to hypoxia, whereas they became silent under normoxia (refer to Figs. 2~4). These findings strongly suggest that channel conductance can be separately regulated along with channel activity in response to hypoxia.
There have been many studies on K+ channel activity in response to acute hypoxia [4,5,7]. In a study on Kv1.2 and Kv2.1 in PC12 cells, hypoxia inhibited oxygen-sensitive K+ (KO2) current, resulting in membrane depolarization and peak inhibition within 2 min, which correlated well with the time course of membrane depolarization. The amplitude of depolarization depended on the grade of hypoxia, i.e. level of low PO2 [6]. However, it was also revealed that exposure to hypoxia for more than 2 min caused further depolarization (see details in Fig. 2A and 2B in reference 6). For persistent hypoxia, membrane depolarization could continue due to the influx of cations through another channel in addition to the closure of KO2 channels. In this context, 30-pS Na+ channels are proper since they are activated at least 2~3 min following exposure to hypoxia, facilitating further depolarization.
The main focus of this study was to identify a channel other than a KO2 channel activated by acute hypoxia. Our study was performed in PC12 cells using solution half saturated with oxygen (91 mmHg O2) in normal solution (152 mmHg O2). This is a disadvantage of our study compared to those performed under almost anoxic conditions (<20% of normal). However, our conditions are more physiological, considering that the O2 level cannot reach an anoxic state within the few minutes of acute ischemic injury. Under these conditions, the present study first identified that the Na+ channel was activated by acute hypoxia and has a single channel conductance of 30 pS with ENaC-like properties. However, it is unclear whether or not this channel protects from hypoxia/ischemia. With respect to hypoxia, further study on the ENaC-like channel can provide therapeutic implications for neurons in response to hypoxia.
 XML Download
XML Download