

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Invertebrates, like vertebrates, possess innate immune defense systems against bacterial, fungal, and viral pathogens [1,2]. Pathogen-associated molecular patterns (PAMPs) such as lipoteichoic acid (LTA), lipopolysaccharide (LPS), mannan, β-1,3-glucan, lipoproteins, and peptidoglycan have molecular sequences that are consistently found on the surfaces of pathogenic organisms [3,4]. PAMPs are recognized by pattern-recognition proteins (PRPs), which are secreted molecules that circulate in the hemolymph of invertebrates that lack acquired immune systems and bind to pathogens, thereby initiating signals leading to the release of pathogens [5,6].
Lectins are carbohydrate-binding proteins of nonimmune origin [7,8], and are important PRPs found in hemolymph along with coagulation factors, protease inhibitors, antimicrobial substances, and other proteins in invertebrates [1,2,9]. Lectins bind to specific carbohydrates on the surfaces of pathogens to activate complementary antipathogenic pathways [9,10].
Numerous studies of the structure of plant lectins and their carbohydrate-binding regions have revealed that they consist mainly of β-structures [11]. However, little information on the structure and function of marine invertebrates has been reported [12].
The marine crab Philyra pisum belongs to the family Leucosiidae and is distributed throughout the muddy areas of the west coast of South Korea. Our previous investigation indicated that P. pisum lectin (PPL), which has a molecular mass of 24,060 Da and is obtained from the hemolymph of P. pisum, exhibits hemagglutination and antiproliferative activities in human lung cancer cells [13]. The molecular characterization of PPL including its molecular mass, pI value, amino acid sequences, amino acid composition, and effects of pH, temperature, and metal ions on its activity as well as mitogenic activities on human lymphocytes were also investigated [14].
In the present study we investigated the characterization of PPL, such as by analyzing its monosaccharide composition, its secondary structure, and the structural effect of glycan, as well as its possibility of binding to different PAMPs and its antiproliferative effects on human cancer cells.
METHODS
Purification and hemagglutination activity
Hemolymph was prepared from P. pisum and the PPL was purified from this hemolymph as described previously [13,14]. The concentration of the purified lectin was determined by the Bradford assay [15] using bovine serum albumin (Bio-Rad, Hercules, CA, USA) as the standard. The hemagglutination activity was assayed using native erythrocytes at 2% according to a previously described method [13,16].
Monosaccharide composition analysis
PPL (0.1 mg) was incubated with 2 M trifluoroacetic acid (Sigma, St. Louis, MO, USA) for 4 h at 100℃ to obtain the neutral and amino sugars. Monosaccharides were separated using high-pH anion-exchange chromatography (HPAEC) with a pulsed amperometric detector (PAD) (Dionex, Sunnyvale, CA, USA) equipped with a CarboPac™ PA10 column (4×250 mm) (Dionex) and an AminoTrap™ column (4×20 mm) (Dionex) working at an isocratic concentration of 20 mM NaOH (Fisher Scientific, Fair Lawn, NJ, USA) and at a flow rate of 0.5 ml/min at room temperature. The concentration of each monosaccharide was quantified from a calibration curve constructed using a standard mixture containing twofold dilutions from 800 pmol of each monosaccharide. The monosaccharides used in this study (e.g., fucose, galactose, glucose, mannose, galactosamine, glucosamine, xylose, and arabinose) were all purchased from Sigma.
Secondary structure
Circular dichroism (CD) spectrum analysis was performed in the far-UV region (190~260 nm) using a spectrophotometer (J-715, Jasco, Tokyo, Japan). The temperature was maintained at 20℃ when analyzing samples (0.1 mg/ml) in 20 mM ammonium bicarbonate (pH 8.0). The scan speed was 100 nm/min, the response time was 4 sec, and the bandwidth was 1.0 nm. The CD spectrum was plotted on a millidegree ellipticity scale.
Preparation of deglycosylated PPL
The PPL glycans were chemically released using trifluoromethanesulfonic acid (TFMS) contained in the Glycoprofile IV chemical deglycosylation kit (Sigma), and the reaction was performed according to the manufacturer's instructions. The deglycosylated PPL was lyophilized and stored at -20℃.
Antiproliferative activity
Human breast cancer MCF-7 and human bladder cancer 5637 cell lines were purchased from Korean Cell Line Bank. Cells (8×103 cells/0.1 ml/well) were incubated with PPL (100 µl) in 96-well culture plates for 72 h in a humidified atmosphere of 5% CO2 at 37℃. The percentage viability of the cancer cells was determined using the 2-(2-methoxy-4-nitrophenyl)-3-(4-nitrophenyl)-5-(2,4-disulfophenyl)-2H-tetrazolium, monosodium salt (WST-8, Dojindo, Kumamoto, Japan) assay according to a modified version of Tominaga's method [17], and quantified as {(absorbance of treated cells)/(absorbance of untreated cells)}×100.
RESULTS
Monosaccharide composition analysis
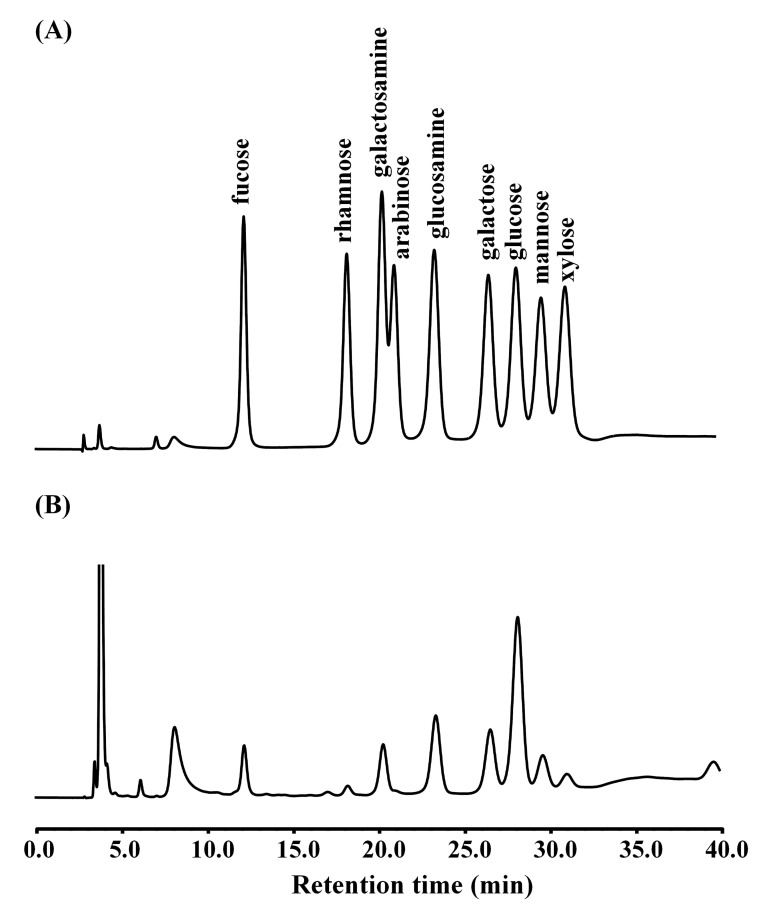
The purified PPL exhibited the same hemagglutination activity as reported previously [13] (data not shown). The composition of each monosaccharide in the purified PPL was identified by comparing the retention time of the monosaccharide peak in the standard obtained using the HPAEC-PAD method (Fig. 1). The monosaccharide ratio was calculated as a percentage based of the moles of monosaccharide per mole of PPL. As shown in Fig. 1, the monosaccharides of PPL (50 pmol) mainly comprised glucosamine (166.8±2.2 pmol, mean±SD; 35.3%), galactose (145.4±0.3 pmol, 30.8%), galactosamine (68.5±0.5 pmol, 14.5%), fucose (54.4±1.3 pmol, 11.5%), and mannose (37.3±1.3 pmol, 7.9%).
Hemagglutination inhibition assay
We investigated the carbohydrate-binding specificity of PPL using a competitive inhibition assay with PAMPs at various concentrations. As indicated in Table 1, the hemagglutination activity of PPL was inhibited by all kinds of LTAs from the various Gram-positive bacteria used in this study, including those from Staphylococcus aureus (8 µg/ml), Streptococcus pyogenes (31 µg/ml), Streptococcus faecalis (4 µg/ml), and Bacillus subtilis (2 µg/ml), as well as by mannan from Saccharomyces cerevisiae (31 µg/ml). However, LPSs from Escherichia coli 055:B5, E. coli 0111:B4, and Klebsiella pneumoniae or laminarin from Laminaria digitata did not lead to inhibition even when they were applied at concentrations up to 500 µg/ml.
Secondary structure
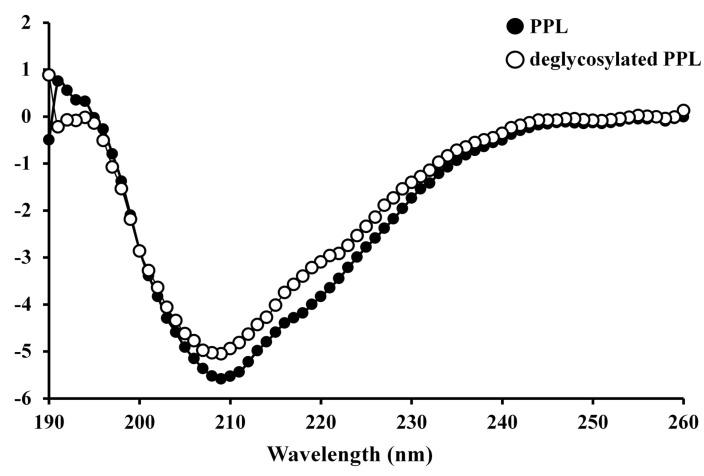
The CD spectrum of PPL in its native form was characterized by minima at 209 nm and 217 nm and a maximum at 191 nm, with a negative-to-positive crossover at 194 nm, which manifested as ellipticity (Fig. 2). The secondary structure of PPL was determined using the protocol reported by Yang et al. [18], which revealed that its estimated α-helix, β-sheet, β-turn, and random coil contents were 14.0%, 39.6%, 15.8%, and 30.6%, respectively; the corresponding contents of these structures in deglycosylated PPL were calculated as 13.7%, 40.0%, 15.8%, and 30.5%, respectively.
Antiproliferative activity
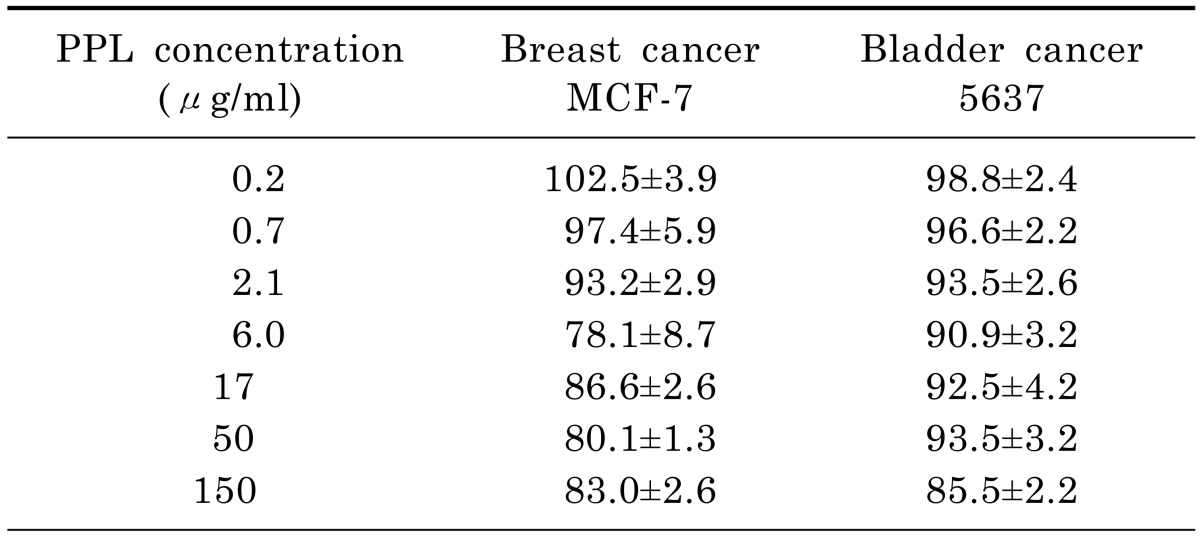
The antiproliferation against human breast and bladder cancer cells was slightly decreased, but the percentage viabilities for both types of cell still exceeded 80% of that for the untreated control cells. Accordingly, PPL exerted no significant antiproliferative effects on human breast cancer MCF-7 cells or human bladder 5637cells (Table 2).
DISCUSSION
The results from the hemagglutination inhibition assays indicated that PPL binds with high specificity to both LTA and mannan-despite them sharing no common structures-but not to LPS or β-1,3-glucan. LTA and LPS are the cell wall components of Gram-positive and Gram-negative bacteria, respectively, while mannan and β-1,3-glucan are constituents of fungal cell walls [4,19,20].
There have been reports of a mannose (mannan)-binding lectin that specifically binds to bacterial LTA through either a carbohydrate recognition domain or another part of the domain [21,22]. The present results showing differential binding to microbial components such as LTA or mannan indicated that different regions of PPL probably participate in binding to microbial components in a similar way to previously reported lectins [23]. Furthermore, our data show that recognition is the initiation process in various immune responses in this crab species, such as prophenoloxidase activation, phagocytosis, nodule formation, and encapsulation [2,3].
The monosaccharide analysis revealed that PPL is a glycoprotein composed of N-acetylglucosamine, galactose, N-acetylgalactosamine, fucose, and mannose, since of de-N-acetylation of monosaccharides such as N-acetylgalactosamine and N-acetylglucosamine occurred during acid hydrolysis. Studies investigating the effect of glycosylation on glycoprotein structure have mainly used completely deglycosylated proteins generated through chemical degradation with reagents such as TFMS [24]. However, the present study has demonstrated that the secondary-structure contents of deglycosylated PPL are quite similar to those of intact PPL, mean that glycans exert almost no effect on the intact structure of PPL.
The proportions of α-helix (4~61%) and β-sheet (6~57%) structures vary widely in marine invertebrate lectin [25-29]. In contrast, the PAMP-binding domain of PRP obtained from the hemolymph of a pyralid moth reportedly consists primarily of α-helix, with only a small amount of β-sheet [30]. PPL contains a relatively low proportion of α-helix, which is broadly consistent with reports for legume lectins (as also determined using the protocol of Yang et al.) including a low proportion (less than 30%) of α-helix and a high proportion (more than 60%) of β-sheet [11,31,32].
There have been numerous reports on the structure and carbohydrate-binding specificity of plant lectins. However, little information on lectins isolated from marine invertebrates has been reported. Many lectins exert antiproliferative effects against human cancer cells, but the mechanisms of action are not fully understood [33]. These antiproliferative activities are probably related to the type and linkage of the glycoconjugate on the surface membranes of cancer cells [8].
In conclusion, the present results indicate that PPL is a glycoprotein whose secondary structure includes a relatively low proportion (14.0%) of α-helix and a high proportion (55.4%) of β-structure, and that this is not affected by glycans. PPL binds to both bacterial LTA and fungal mannan. Our previous study showed that PPL exerts antiproliferative effects on human lung cancer cells [13]; in contrast, the present study found no such effects on human breast and bladder cancer cells. These properties are inherent to the innate immune system of P. pisum and indicate that PPL could be involved in signal transmission into Gram-positive bacteria or fungi. Further investigations including a detailed structural analysis and determination of the binding constant to LTA or mannan as well as the binding sites to PAMPs of PPL are underway in our laboratory.

 XML Download
XML Download