

 PDF
PDF ePub
ePub Citation
Citation Print
Print


ABBREVIATIONS
MDR3
multidrug resistance 3
LPAC
low phospholipid-associated cholelithiasis
ABCB4
ATP-binding cassette, subfamily B, member 4 gene
PFIC3
progressive familial intrahepatic cholestasis 3
UDCA
ursodeoxycholic acid
SNP
single nucleotide polymorphism
TMs
transmembranes
ICDs
intracellular domains
ECs
extracellular loops
TMD-NBD
transmembrane domain-nucleotide binding domain
INTRODUCTION
Human multidrug resistance 3 (MDR3) coded by ATP-binding cassette, subfamily B, member 4 gene (ABCB4) belongs to ABC transporters family [1]. MDR3 is expressed on the canalicular membrane of the hepatocyte and translocates phosphatidylcholine from the inner to the outer leaflet of the canalicular membrane, playing an important role in protection of the liver from bile acids [2]. It was known that dysfunction of MDR3 due to genetic variations can cause several hepatic diseases in human, such as low phospholipid-associated cholelithiasis (LPAC), progressive familial intrahepatic cholestasis 3 (PFIC3), obstetric cholestasis, and drug-induced liver injury [3-7]. LPAC is intrahepatic cholelithiasis characterized by at least two of the following criteria: age onset of symptom less than 40 years, intrahepatic echogenic foci or microlithiasis, and recurrence after cholecystectomy [8,9]. LPAC is occurred higher in female than male, while its exact prevalence rate is not known. LPAC patients often have to undergo cholecystectomy due to biliary pain which could not be relieved by the treatment using ursodeoxycholic acid (UDCA). In addition, a few patients with end-stage liver disease can be candidate for liver transplantation [8].
To date, there were few studies to investigate the expression or function of each ABCB4 variant in spite of the clinical importance of MDR3 transporter. Most previous functional analysis of ABCB4 genetic variations have been restricted to missense mutations identified in PFIC3 patients. For example, a ABCB4 mutation found in PFIC3 patients, I541F, was shown to decrease transport activity through reduction of membrane MDR3 expression [10,11]. Recently, we identified and functionally characterized the ABCB4 promoter variants through direct sequencing using genomic DNA samples from 126 Koreans and reported 2 common promoter haplotypes of ABCB4 resulted in significantly decreased promoter activity [12].
Previously, mutation analysis of ABCB4 was performed using genomic DNA samples from 32 LPAC patients [3]. The authors identified 14 mutations in the coding region including 9 missense and 5 nonsense mutations. They found that all these mutations were not present in other two groups: one was group consisting patients with a classic gallstone disease and the other was group of patients without a history of cholelithiasis. However, a functional characterization of each mutant was not performed in their study.
In this study, we selected 3 novel missense mutations of ABCB4 that were first reported by Rosmorduc et al. [3] and investigated the function of each mutant using various in vitro assays such as membrane vesicular transport, immunoblotting, and surface protein biotinylation. To our knowledge, this is the first study to characterize functionally ABCB4 mutations found in LPAC. This study may contribute to the development of diagnostic kits for LPAC in the future.
METHODS
Construction of ABCB4 plasmids
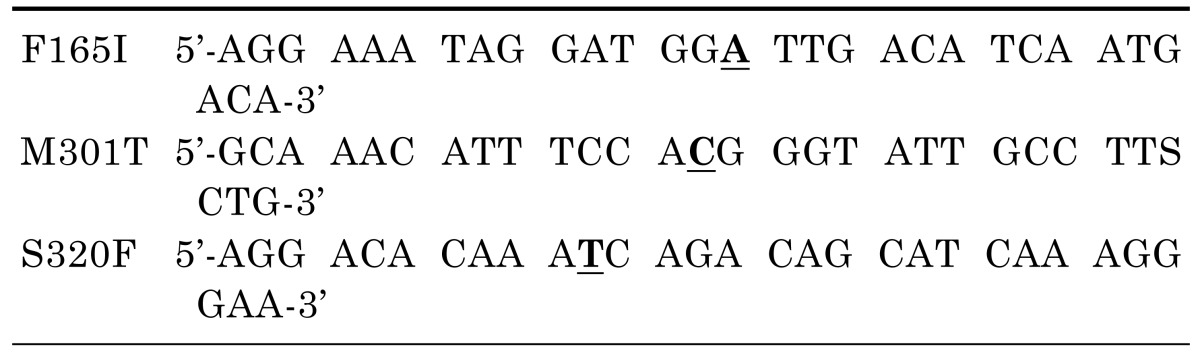
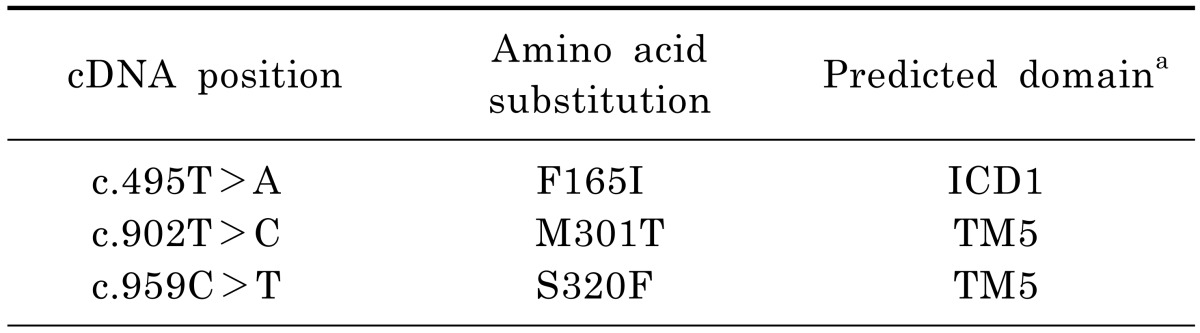
To construct the plasmid containing a reference ABCB4 gene, vector (BC_042531) was purchased (Thermo Fisher Scientific Inc., Waltham, MA, USA) and subcloned into the pcDNA3.1(+) vector. Plasmids containing the mutant ABCB4 sequences were produced using QuikChange® II Site-Directed Mutagenesis Kit (Agilent Technologies, Santa Clara, CA, USA) with the primers listed in Table 1 from the pcDNA3.1-ABCB4 plasmid. Nucleotide location numbers were assigned from the translational start site according to the ABCB4 mRNA sequence (GenBank accession number; NM_018849.2).
Membrane vesicle preparation
Preparation of membrane vesicles was performed according to a previously described method [13]. Briefly, ABCB4 reference or mutant-bearing plasmids were transfected into HEK-293T (Human embryonic kidney) cells using the Calcium Phosphate Transfection Kit (Life Technologies Corporation, Carlsbad, CA, USA). Cells were harvested 48 h later in a homogenization buffer supplemented with a protease inhibitor cocktail. Harvested cells were then subjected to nitrogen cavitation at 350 pounds per square inch for 15 min and transferred to a tube containing 0.5 M EDTA. The membrane vesicle fractions were collected by sucrose density gradient centrifugation at 1,000,000×g for 90 min. Immediately after preparation, the vesicles were suspended in buffer containing 250 mM sucrose and 50 mM Tris (pH 7.4) at a protein concentration of 4~8 mg/ml.
Adenosine triphosphatase (ATPase) assay for paclitaxel transport
ATPase assays for paclitaxel transport were performed according to a previously described method with some modifications [14]. The vanadate-sensitive ATPase activity was measured using the SensoLyte® MG Phosphate Assay Kit (71103, AnaSpec, Fremont, CA, USA), to assess the paclitaxel transporting capacity of MDR3. Briefly, membrane vesicles (4 µg/well) were incubated in 4 mM MgCl2, 5 mM 3-(Nmorpholino)-propanesulfonic acid-Tris (pH 7.0), 4 mM ATP, and various concentrations of paclitaxel with or without 1 mM sodium orthovanadate at 37℃ for 15 min. Finally, absorbance was measured at 620 nm using a microplate reader. ATPase activities were determined as the difference in the inorganic phosphate liberation in the presence or absence of sodium orthovanadate.
Immunoblotting
The ABCB4 reference or mutant-bearing plasmids were transfected into HEK-293T cells using the Lipofectamine LTX and Plus reagents (Life Technologies). 48 h after transfection, immunoblotting was performed using the following primary antibodies: a mouse anti-MDR3 antibody (P3II26, Abcam, Cambridge, UK), rabbit anti-neomycin phosphotransferase II antibody (06-747, Millipore, Billerica, MA, USA), or goat anti-β-actin antibody (sc-1616, Santa Cruz Biotechnology, Santa Cruz, CA, USA). The intensity of each band was measured using ImageJ (National Institutes of Health, Bethesda, MD, USA).
Biotinylation of cell surface proteins
Biotinylation experiments were conducted using a Cell Surface Protein Isolation Kit (Thermo Fisher Scientific Inc.) according to the manufacturer's protocol using the HEK-293T cells obtaining from transfection of the ABCB4 reference or mutant-bearing plasmids. A rabbit polyclonal anti-Na+/K+ ATPase α-1 antibody (06-520, Millipore) was used as an internal standard.
Immunofluorescence
For immunofluorescence, HEK-293T cells were grown on coverslips in a 24-well plate and the reference or mutant ABCB4-bearing plasmids were transfected into HEK-293T cells. 48 h after transfection, cells were incubated with CellMask Plasma Membrane stain solution (Life Technologies) and washed 3 times. After fixation with 4% PFA, cells were permeabilized with acetone and blocking was performed. For detection of MDR3, the anti-MDR3 antibody and Alexa Fluor® 488 rabbit anti-mouse IgG (11059, Life Technology) was used. Confocal images were captured using confocal laser scanning microscope (LSM5 PASCAL; Carl Zeiss, Oberkochen, Germany) and the digital images were analyzed using a LSM Image Examiner (Carl Zeiss).
Statistical analysis
Statistical analysis was performed using GraphPad Prism 4.0 software package (GraphPad Software, Inc., San Diego, CA, USA). All p values were calculated using oneway analysis of variance followed by Dunnett's two-tailed test, and p values less than 0.05 were considered to be statistically significant.
RESULTS
Mutations of ABCB4 examined in this study
To perform a molecular characterization of ABCB4 mutations found in LPAC patients, we selected some novel mutations that were reported by Rosmorduc et al. [3]. They identified 14 ABCB4 mutations including 9 missense mutations through direct sequencing of genomic DNA samples from 32 LPAC patients. Among these mutations, we excluded mutations that resulted in protein truncation or whose effect on other ABC transporters had already been investigated. Table 2 shows the 3 missense ABCB4 mutations included in this study. It has known that MDR3 consists of 12 transmembranes (TMs), 6 intracellular domains (ICDs), 6 extracellular loops (ECs), and a linker connecting the N-terminal to the C-terminal transmembrane domain-nucleotide binding domain (TMD-NBD) [15]. The ABCB4 mutant, F165I might be located in the ICD1, while other two mutants, M301T and S320F are located in the TM5 [3].
The effect of mutations on the transport activity of MDR3
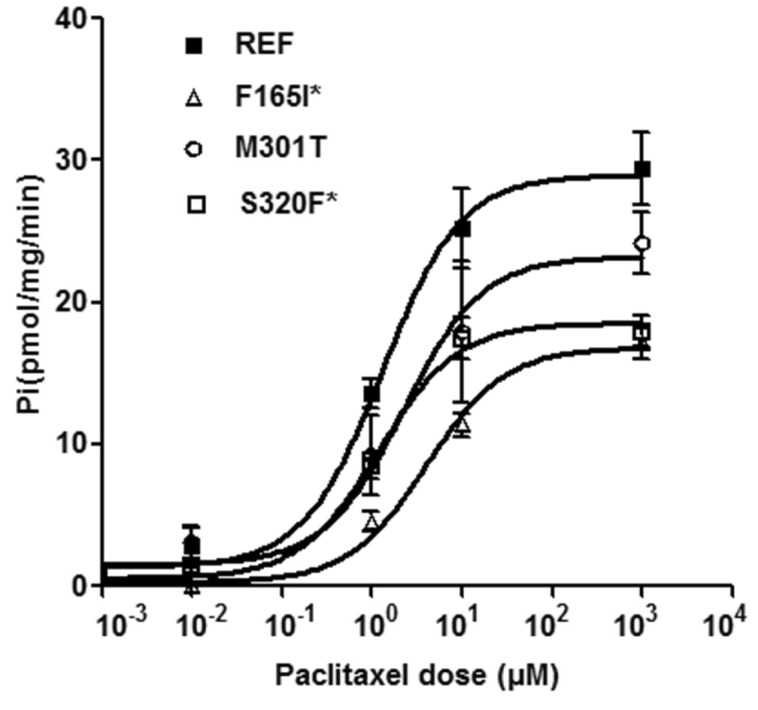
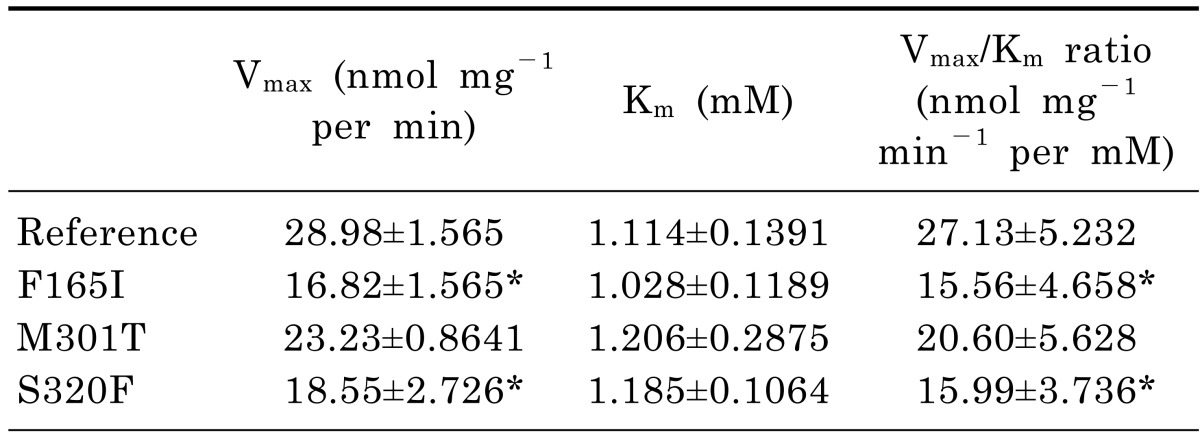
It has been previously shown that MDR3 transports paclitaxel [16]. To characterize the functional effects of the ABCB4 mutations, we first measured the ability of each mutant to transport paclitaxel. Fig. 1 shows the results of the ATP-dependent paclitaxel uptake in inside-out membrane vesicles expressing ABCB4 reference or mutations at various concentrations (0.01, 1, 10, and 1,000 µM) of paclitaxel. To exclude ATPase activity by other endogenous ABC transporters including MDR1, values for transport activity were obtained by subtracting the uptake in empty vector-transfected cells from that in cells transfected with ABCB4 reference or mutant-bearing vectors, at each corresponding paclitaxel concentration. The uptake of paclitaxel was significantly reduced in membrane vesicles expressing 2 ABCB4 mutations, F165I and S320F. M301T mutant also showed a reduction in the transport activity compared to that of the reference, although statistical significance was not observed. The paclitaxel Vmax and Km values for the ABCB4 reference or mutant genes are shown in Table 3. We observed that the average values of Vmax/Km for F165I and S320F were significantly reduced compared to that of the ABCB4 reference. This was attributable to a dramatically reduced Vmax. In the case of Km, these mutants showed similar Km values compared to that of the reference.
The effect of mutations on the expression level of MDR3
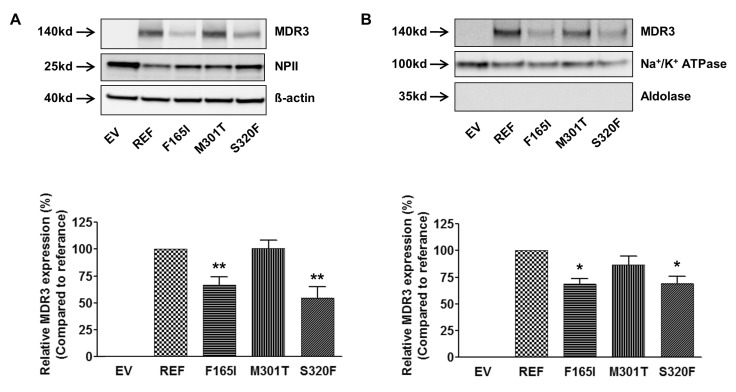
Cell surface biotinylation and immunoblotting were performed to investigate the mechanisms through which ABCB4 mutations alter the transport activity. Fig. 2A shows the results from immunoblotting of the total cell lysates after transfection of ABCB4 reference or mutant-bearing plasmids into HEK-293T cells. The 2 mutations, F165I and S320F that showed decreased transport activities as compared to the reference, had significantly decreased MDR3 expression as compared to reference. The MDR3 expression level of M301T was comparable with that of the reference. Then, we investigated MDR3 expression levels of these mutants on the plasma membrane by cell surface biotinylation and observed that those of F165I and S320F were significantly decreased by 31%, compared to that of the reference (Fig. 2B). The results from immunoblotting or cell surface biotinylation experiments could explain the reason of altered transport activities of ABCB4 mutants, F165I and S320F; the decreased transport activities of these mutants were due to the reduced expression of functional MDR3 on the plasma membrane.
The effect of mutations on the subcellular localization of MDR3
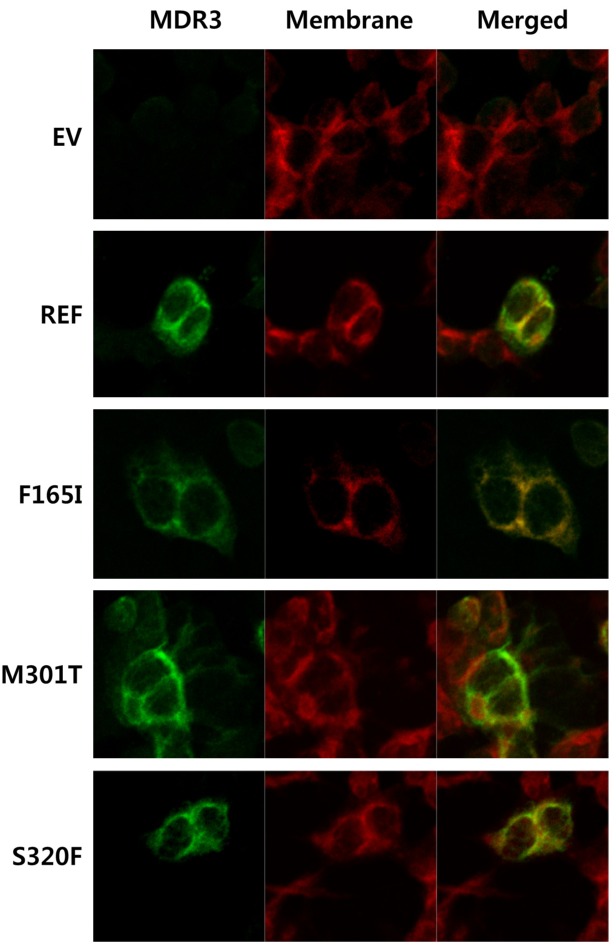
To investigate the subcellular localization of MDR3 and its mutants, immunofluorescence was performed after transfection of ABCB4 reference or mutant-bearing plasmids into HEK-293T cells. Fluorescence microscope showed that the majority of the reference MDR3 was restricted to the plasma membrane and was co-localized with the plasma membrane staining (Fig. 3). The subcellular expression of M301T was comparable with that of the reference while the co-localization of F165I and S320F with plasma membrane was decreased. These findings were consistent with the results obtained in immunoblotting and cell surface biotinylation experiments.
DISCUSSION
MDR3 is highly expressed on the canalicular membrane of hepatocytes and mediates the excretion of bile acids [1]. Functional changes in the activity or expression of MDR3 caused by genetic variants have been implicated as the primary genetic risk factor of LPAC.
Previously, mutation analysis of ABCB4 was performed using genomic DNA samples from 32 patients with LPAC [3]. The authors identified 14 mutations in the coding region including 9 missense and 5 nonsense mutations that can be considered as potential disease-causing mutations. However, the functional characterization of each mutant was not performed in their study.
The current study was conducted to characterize functionally the ABCB4 mutations found previously by Rosmorduc et al. [3]. Among these 14 ABCB4 mutations, we selected 3 mutations after excluding mutations that were nonsense mutations or had already been investigated in relation to other ABC transporters. Through a membrane vesicular transport assay using paclitaxel, we found that 2 mutants, F165I and S320F, showed significantly decreased transport activity compared to the reference (Fig. 1). F165I and S320F are located in the ICD1 and TM5 of MDR3, respectively, that might be involved in coupling the energy from ATP hydrolysis to substrate transport and the conformational change involved in substrate extrusion, respectively [3,15]. M301T, showing a comparable transport activity with that of the reference is also located in TM5. The different effect of genetic variations which are located closely on the transport activity was also reported previously [17]. These data suggest that the function of each variation could not be determined only by its location on the transporter. The reduced transport function of F165I and S320F mutants also might be due to the decreased expression of functional MDR3 on the plasma membrane (Fig. 2). In the kinetic studies using paclitaxel, we observed that all Vmax values of the mutants showing decreased transport activity were significantly decreased compared to the reference, whereas the changes in the Km values were not statistically significant (Table 3). These data support that the reduced transport activity of these mutants is primarily attributable to the reduced expression of functional MDR3 on the plasma membrane.
One of the ABCB4 mutations, I541F, was found in PFIC3 patients and was shown to be a trafficking-defective mutation [10,11]. This mutant demonstrated decreased transport activity due to the marked reduction of membrane MDR3 expression. 2 mutants, F165I and S320F, examined in this study also showed decreased transport activity and protein expression, although the extent of reduction was less than that of I541F. Therefore, PFIC3 patients are usually candidates for liver transplantation before adulthood due to complications such as hepatic failure, liver cirrhosis, and hepatocellular carcinoma, while the majority of LPAC patients can be controlled by UDCA if this treatment is started early [8,18].
According to the previous study by Rosmorduc et al. [3], only 18 patients had ABCB4 mutations among the 32 patients clinically diagnosed as LPAC. As mentioned above, UDCA treatment should be started early to control LPAC effectively. Therefore, it would be valuable to develop the diagnostic kits for the rapid detection of ABCB4 mutations.
In this study, we could not assess the correlation between the severity of clinical symptoms and the molecular function of mutants because the previous study that was referred in this study [3] did not provide the clinical symptoms and mutant type of each patient. Recently, screening of genomic DNA samples from 156 LPAC patients identified 46 ABCB4 mutations, consisting of 13 nonsense and 33 missense mutations. Among the 33 missense mutations, 15 were novel [9]. By investigating the genotype-phenotype relationships in LPAC patients, the authors found that the nonsense mutations were associated with an earlier onset of symptoms, while biliary complications or the frequency and severity of intrahepatic cholestasis of pregnancy were not related with the mutant type. However they didn't perform in vitro assay of each mutant. To clarify these relationships further, the functional characterization of each mutant would be necessary.
In conclusion, we characterized the ABCB4 mutations found in LPAC patients and revealed that 2 mutants showed significantly decreased transport activity, mainly due to decreased functional MDR3 expression on the cell membrane. To our knowledge, this is the first study to characterize functionally ABCB4 mutations that cause LPAC. This study may contribute to the development of diagnostic kits for LPAC in the future.
 XML Download
XML Download