

 PDF
PDF ePub
ePub Citation
Citation Print
Print


ABBREVIATIONS
RANKL
receptor activator of NF-κB ligand
NFATc1
nuclear factor of activated T cells cytoplasmic 1
AlF4-
aluminum fluoride
BMMs
bone marrow-derived macrophages
TRAP
tartrate-resistant acid phosphatase
MAPK
mitogen-activated protein kinase
FcRγ
Fc receptor common γ subunit
ROS
reactive oxygen species
OGR1
ovarian cancer G protein-coupled receptor 1
GAP
GTPase activating protein
MNCs
multinucleated cells
RGS
regulator of G-protein signaling
CREB
cAMP response element binding proten.
INTRODUCTION
Receptor activator of NF-κB ligand (RANKL) expressed from osteoblasts binds to its own receptor, RANK, in osteoclast precursor cells, bone marrow-derived macrophage (BMMs), and initiates osteoclastogenesis by activating various intracellular signal pathways including mitogen-activated protein kinases (MAPKs), NF-κB, AP-1, c-fos, and NFATc1 [1-5]. Among them, NFATc1, which is regarded as a key factor to determines the late-stage of differentiation to osteoclast, is well defined to be modulated by intracellular Ca2+ mobilization [3]. According to previous reports including ours, RANKL generates intracellular Ca2+ mobilization via co-stimulatory signals mediated through immunoreceptor tyrosine-based activation motif (ITAM)-harbouring adaptors, such as Fc receptor common γ subunit (FcRγ) and DNAX-activating protein (DAP) 12 and reactive oxygen species (ROS) generation. RANKL-mediated intracellular Ca2+ mobilization is presented in a form of oscillations which needs Ca2+ flux into cytoplasm from both external and internal Ca2+ stores to form and sustain oscillation frequencies that is essential for sequential activation of Ca2+/calmodulin-dependent kinase, calcineurin, and NFATc1 [6,7]. In contrast, it has not been reported that Ca2+ mobilization generated by RANKL-independent way affects on differentiation into osteoclast.
Diverse signal pathways mediated by G-protein coupled receptors (GPCR) is reported to be involved in various osteogenic activities including cell survival, tumorigenesis, and differentiation of osteoclast [8-12]. For example, ovarian cancer G protein-coupled receptor 1 (OGR1) activated by protons or lysolipids modulates not only osteoclast survival through NFAT-independent but also osteoclastogenesis through an OGR1/NFAT pathway [8,11]. Notably, regulator of G-protein signaling (RGS) 18, which is known to act as a GTPase activating protein (GAP), negatively regulates osteoclastogenesis by modulating the activity of Gα subunit [11]. Here an important question has arisen whether modulating the activity of Gα subunit alone affects on the RANKL-induced Ca2+ oscillations and osteoclastogenesis.
Aluminum-fluoride complex (AlF4-) act as an analog of a phosphate group and stimulates cellular heteromeric G-proteins because of its structural similarity with phosphate group. AlF4- is tetrahedral and its Al-F bond length is very similar to P-O bond length of phosphate [13]. AlF4- can be used as useful tools investigating signal pathways following G-proteins. AlF4- stimulates G-protein and mimics the action of many neurotransmitters, hormones, and immune system [13,14]. AlF4--induced Ca2+ oscillations were showed in smooth muscle cell [15] and pancreatic acinar cells [16]. AlF4- is also known to transmit signals modulating activities of bone cells, such as cell proliferation, differentiation [17] and protein phosphorylation [18]. It was reported that the effects of fluoride and aluminum on levels of the second messenger molecules are dependent on the type of cells and tissues [13]. Along with these reports, we postulated that induced Ca2+ signaling by aluminum-fluoride complexes may affect osteoclast differentiation, and the investigation was undertaken to study the effects of AlF4- on Ca2+ signaling and osteoclasts differentiation in primary cultured mouse bone marrow-derived macrophages (BMMs). In this study, we demonstrate that co-stimulation of AlF4- with RANKL has synergistic effects enhancing RANKL-induced Ca2+ oscillations, NFATc1 expression, and forming multinucleated cells (MNCs).
Go to : 
METHODS
Chemicals and antibodies
Recombinant mouse soluble RANK ligand and recombinant mouse M-CSF were purchased from KOMA Biotech (Seoul, Korea). AlCl3 and NaF were purchased from Sigma Aldrich (St. Louis, MO, USA) and Fluka (Buchs, Switzerland) respectively. Fura-2/AM was purchased from Teflabs (Austin, TX, USA). Pluronic F-127 was obtained from Invitrogen (Eugene, Oregon, USA). Monoclonal antibodies against ERK, phospho-ERK, JNK and phospho-JNK were purchased from Cell Signaling (Danvers, MA, USA); polyclonal antibody for NFATc1 was from Santa Cruz Biotechnology (Santa Cruz, CA, USA).
Cell culture
Non-adherent bone marrow-derived macrophages (BMMs) were isolated from tibia and femur of 4-week-old male ICR mice (weight 21~25 g) as described previously [19]. Briefly, isolated BMMs were cultured in α-minimum essential medium (α-MEM; Gibco/BRL, Grand Island, NY, USA) supplemented with 10% fetal bovine serum (FBS; Gibco/BRL, Grand Island, NY, USA) in the presence of M-CSF (20 ng/ml).
Intracellular Ca2+ imaging
The cells were seeded on cover glass in a 35-mm dish (1×105 per dish) and treated with RANKL (50 ng/ml) and/or aluminum-fluoride complex (1.25 mM NaF+25 nM AlCl3). Cells were loaded with 3 µM fura-2/AM, 0.05% Pluronic F-127 in an extracellular physiological salt solution (140 mM NaCl, 5 mM KCl, 1 mM MgCl2, 1 mM CaCl2, 10 mM HEPES, and 10 mM glucose, titrated to pH 7.4 with NaOH. The osmolarity of the PSS was 310 mOsm) at room temperature and washed out with standard solution. [Ca2+]i was measured using wave lengths of 340 and 380 nm, and the emitted light was passed through a 510 nm cut off filter and was collected with a CCD camera and analyzed with a Meta Fluor system (Universal Imaging Co., Downingtown, PA, USA). The 340/380 fura-2 ratio was taken as a measure of [Ca2+]i and fluorescence images were obtained at 2s intervals. Images were digitized and analyzed through MetaFluor software (Universal Imaging).
In vitro osteoclastogenesis and TRAP staining
RANKL (50 ng/ml) and/or AlF4- (1.25 mM NaF+25 nM AlCl3) were added to BMMs (2×105 per well) cultured in 24-well plate with α-MEM containing M-CSF (20 ng/ml). Six days later osteoclastogenesis was confirmed by TRAP (tartrate-resistant acid phosphatase) staining to evaluate TRAP-positive multinucleated osteoclast formation. Naphthol AS-MX (Amresco, Cleveland, OH, USA) was used as the substrate and fast red violet LB salt (Sigma Aldrich, St. Louis, MO, USA) as the diazonium salt. TRAP+ multinucleated cells (≥3 nuclei) were counted.
Immunocytochemistry
Cells seeded on 12-mm cover glasses were treated with RANKL (50 ng/ml) and/or AlF4- (1.25 mM NaF+25 nM AlCl3). After fixation with 10% methanol at -20℃ for 10 min, cells were washed with cold phosphate buffered saline (PBS) and neutralized with 50 mM Glycine for 10 min at 4℃. Cells were sequentially blocked with 5% goat serum in incubation buffer (0.1% gelatin, 1% BSA, 0.01% sodium azide in PBS) and stained with 1 drop of 200 U/ml phalloidin in the dark for 1 hr. And then, cells were washed with cold PBS.
Western blot
Whole cell extracts of BMMs were prepared by washing the cells with cold PBS and lysed in RIPA buffer (20 mM Tris, pH 7.4, 250 mM NaCl, 2 mM EDTA, pH 8.0, 0.1% Triton-X100, 0.01 mg/ml aprotinin, 0.005 mg/ml leupeptin, 0.4 mM PMSF, and 4 mM NaVO4) for 30 min. 20 µg/well proteins were loaded on 10% SDS-PAGE respectively, and then were separated by size. Separated proteins were transferred onto nitrocellulose membranes, blocked with 5% BSA in TBS, and probed with Abs against phospho-ERK (1:1,000), total-ERK (1:1,000), phospho-JNK (1:1,000), total-JNK (1:1,000), NFATc1 (1:1,000) and β-actin (1: 1,000) for overnight. After washing out, the blots were incubated with HRP-conjugated secondary antibodies for 1 hr, and visualized by enhanced chemiluminescence (ECL, Amersham Pharmacia Biotech, IL, USA) using Agfa CP-BU new film (Mortsel, Belgium). The level of protein expression was digitized film images and analyzed with MetaMorph software (Universal Imaging).
Statistical analysis
Results are expressed as means±S.E. from at least 3 independent experiments. The statistical significances of differences between groups were determined using the Student t-test. The difference is significant if the value is <0.05.
Go to :
RESULTS
Treatment of AlF4- induces Ca2+ mobilization similar to RANKL-induced [Ca2+]i oscillation
We have previously reported that RANKL stimulation on BMMs induces [Ca2+]i oscillations which is dependent on Ca2+ flux from both external and internal store [6]. To determine whether direct activation of G-protein by AlF4--would affect on the intracellular Ca2+ mobilizations, we analyzed AlF4--mediated Ca2+ responses in the presence or absence of RANKL. As shown in Fig. 1, [Ca2+]i oscillations, which is dependent on extracellular Ca2+ influx, were observed from 24 to 48 hrs after RANKL treatment in BMMs (Fig. 1, upper panel). Notably, co-stimulation with AlF4- and RANKL (Fig. 1, middle panel) and AlF4- treatment alone (Fig. 1, lower panel) led to increase of oscillations frequency in comparison to RANKL alone, suggesting that G-protein activation would amplify the RANKL-mediated signals and be enough to induce [Ca2+]i oscillations regardless of the presence of RANKL. Furthermore, these all [Ca2+]i oscillations presented dependency on extracellular Ca2+ influx. Along with these results, we could assumed that [Ca2+]i oscillations caused by AlF4- has similar characteristics with the one caused by RANKL. Moreover, AlF4- is capable to not only induce [Ca2+]i oscillations which has similar characteristics with RANKL-induced [Ca2+]i oscillations but also enhance it.
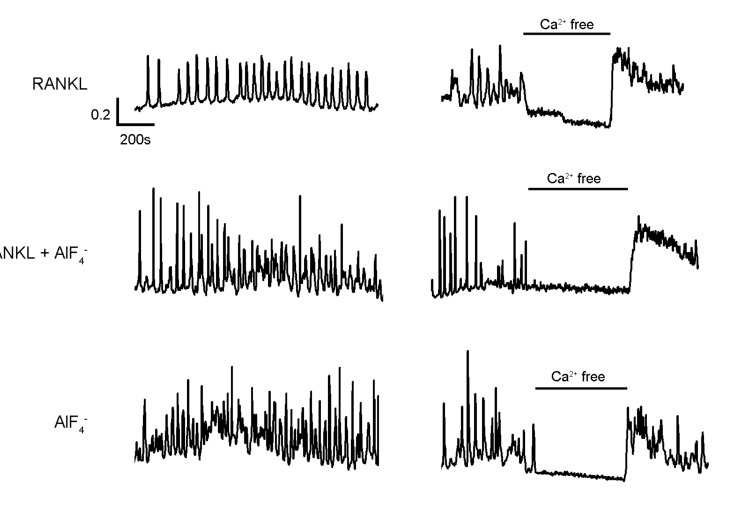 | Fig. 1Effects of the [Ca2+]i increases mediated by RANKL and AlF4- in BMMs. BMMs were treated with RANKL (50 ng/ml) and/or AlF4- (1.25 mM NaF+25 nM AlCl3). After 48 h of the stimulations, [Ca2+]i oscillations in BMMs were measured using Fura-2 fluorescence dye. Compared to RANKL alone, application of AlF4- similarly increased frequency of the [Ca2+]i oscillations in the presence or absence of RANKL (left panels). These induction of [Ca2+]i oscillations were diminished by the removal of extracellular Ca2+ (right panel).
|
Co-stimulation with AlF4- and RANKL enhances NFATc1 expression, but not AlF4- alone
NFATc1 is known to be up-regulated by RANKL stimulation and modulates cell fusion and maturation [3]. As described previously, we found that AlF4- solely induces intracellular Ca2+ mobilization in a form of oscillations which has similar characteristics with RANKL-induced [Ca2+]i oscillations. However, it is not clear that AlF4--mediated [Ca2+]i oscillations is sufficient to transmit signals related on the differentiation to osteoclasts. To answer this question, we employed a standard in vitro osteoclast culture system and prepared whole cell lysates from cells maintained with M-CSF and RANKL in the presence or absence of AlF4-. As shown in Fig. 2, similar tendency of the NFATc1 induction was observed in both samples treated with RANKL alone and both of RANKL and AlF4-, which starts to be increased from 24 hrs and maximized around 48 hrs after stimulation. Whereas, in terms of quantity of expression, co-stimulation with RANKL and AlF4- more than 48 hrs resulted in 1.7-fold increase compared to RANKL alone. Unlike our expectation, sole treatment of AlF4- did not induce NFATc1 expression.
 | Fig. 2Increased expression of NFATc1 in BMMs induced by RANKL and AlF4-. Whole cells lysates were collected from cells stimulated with RANKL and/or AlF4- for indicated time. NFATc1 and actin were blotted with its antibody (upper panels) and then showed means of protein expression levels between RANKL and AlF4- (lower panels). Expression level was significantly increased for NFATc1 in 48 h and 72 h of RANKL and AlF4- treatments (n=3). Data were expressed as the mean±SEM. **p<0.01, ***p<0.001 compared with RANKL treated group.
|
Co-stimulation with AlF4- and RANKL enhances the formation of TRAP-positive multinucleated cells, but not AlF4- alone
To further investigate the effects of AlF4- on RANKL-induced osteoclastogenesis, we examined the effects of enhanced [Ca2+]i oscillations by AlF4- on the formation of MNCs and actin ring. BMMs were stimulated with AlF4- (1.25 mM NaF+25 nM AlCl3) in the presence or absence of RANKL (50 ng/ml) for 6 days, and TRAP and phalloidin staining were conducted. Interestingly, TRAP+ MNCs were only confirmed in RANKL treated cells, but not in the cells with only AlF4- (Fig. 3A). However, BMMs treated with AlF4- and RANKL simultaneously showed remarkably enhanced MNCs formation (Fig. 3A). Consistent with TRAP staining results, we also confirmed that co-stimulation with AlF4- and RANKL increased actin ring formation compared to RANKL alone (Fig. 3B), but not in AlF4- alone. These results suggest that AlF4--mediated signals are not sufficient for cell fusion and attachment but it improves osteoclastogenesis when it coexists with RANKL.
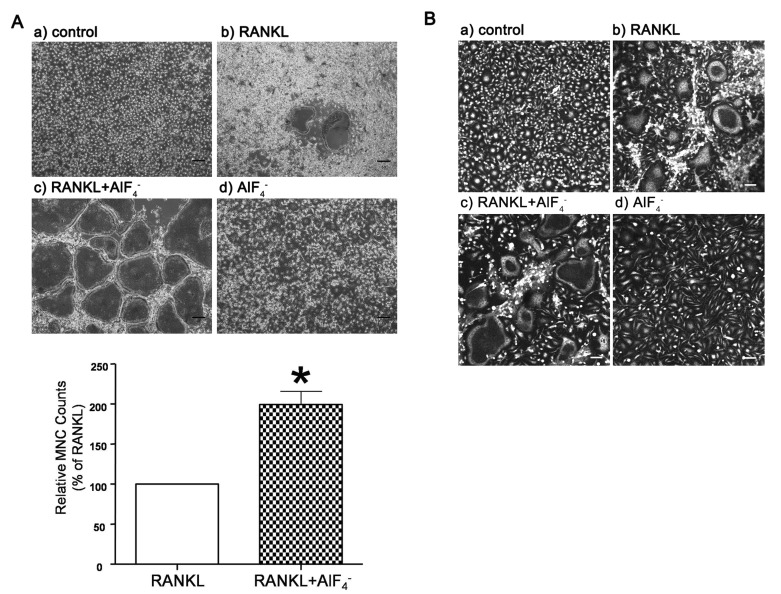 | Fig. 3RANKL and AlF4--induced osteoclast differentiation in BMMs. (A) To confirm the formation of multinucleated cells (MNCs, number of nuclear>3), each sample was incubated for 6 days in (a) control and the presence of (b) RANKL, (c) RANKL and AlF4-, and (d) AlF4-. TRAP staining was performed as described in methods. And then MNCs in each well was counted (n=3, scale bar, 50 µm). Application of RANKL and AlF4- in BMMs was significantly increased rates of TRAP+ MNCs formation more than the application of RANKL alone (lower panel). (B) Actin staining of TRAP+ MNCs in application of RANKL and AlF4- was similarly showed normal actin ring formations during osteoclastogenesis compared to RANKL treated BMM cells. Bar graph shows relative MNCs of BMMs treated with RANKL only and RANKL+ AlF4-. Data were normalized to the number of MNCs in RANKL treated BMMs and expressed as the mean±SEM. *p<0.05 compared with RANKL treated group.
|
Induction of MAPK signal pathway in response to AlF4-
RANKL is also known to transmit signals through activation of MAPK pathway including ERK, JNK, and AP-1 crucial for gene expression by clustering with CREB, MITF, PU.1, and NFATc1 [20-23]. To gain insight of G-protein activation into MAPK pathway, we measured phosphorylation of ERK and JNK in response of RANKL and both of RANKL and AlF4-. To confirm this, western blot analysis was performed on lysates of BMMs incubated with RANKL (50 ng/ml) and/or AlF4- (25 nM AlCl3+1.25 mM NaF) in a time-dependent manner. We could observe phosphorylation of ERK and JNK in response of RANKL, as reported previously, and both of RANKL and AlF4- (Fig. 4). Interestingly, phosphorylation of ERK and JNK was also detected in only AlF4- treated sample, suggesting that activation of G-protein is somehow directly related with MAPK. Taken together, these results clearly indicate that activation of G-protein synergistically improve RANKL-mediated osteoclastogenesis through enhancing Ca2+ oscillations and MAPK pathway.

Go to :
DISCUSSION
Diverse Ca2+ signals in osteoclast are essential for cellular functions including motility, differentiation, and bone resorption [6,8-10]. Several reports including ours clearly presents that RANKL-induced [Ca2+]i oscillation occurred by activation of phospholipase C (PLC) and IP3 production, which evokes Ca2+ release from the ER, resulting in induction of Ca2+ oscillations. There are numerous hormones and neurotransmitters which lead to intracellular Ca2+ mobilization relied on sequential activation of GPCR, G-protein, and PLC [24,25]. Recent studies showed specific GPCRs are involved in osteoclastogenesis and deficiency of RGS impairs to differentiate into osteoclast due to the absence of Ca2+ signaling [26]. Inspired by these, we assumed that activation of G-protein by itself may be sufficient or necessary for the differentiation into osteoclast regardless of RANKL stimulation. Under this hypothesis, we first confirmed that sole activation of G-protein generates [Ca2+]i oscillations using AlCl3 and NaF because traces of aluminum can form aluminum-fluoride complex (most likely AlF4-) easily in aqueous solutions [13] and makes a synergistic action of fluoride to activate G-proteins [27]. AlF4- is well known to directly activate a GTP-binding protein coupled to PLC [28]. Interestingly, we observed that not only co-stimulation with RANKL and AlF4- but also AlF4- alone induce [Ca2+]i oscillations in both BMMs. However, we could not see any Ca2+ responses when the same concentration of AlF4- was treated acutely. Moreover, as our previous report, we also confirmed that extracellular Ca2+ influx is crucial for sustained [Ca2+]i oscillations in response to AlF4- alone and both of RANKL and AlF4-, clearly presenting that AlF4--induced [Ca2+]i oscillations is generated through similar mechanism with RANKL's. The characteristic of intracellular Ca2+ signaling varies as following signal molecules, which gives diversity and versatility on the cell functions [29]. Based on our results, it is assumed that activation of G-protein by AlF4- possibly determines cell fate of BMMs alike with RANKL.
To further investigate physiological phenomenon induced by AlF4- or co-stimulation with RANKL and AlF4-, we decided to check NFATc1 activity and the formation of TRAP+ MNCs and actin ring, which are widely used as a criterion to confirm the osteoclastogenesis. Intriguingly, results from immunoblotting for NFATc1, TRAP staining, and immunostaining for actin revealed that activation of G-protein mediated by AlF4- in the absence of RANKL is not sufficient to differentiate to osteoclast. On the other hand, remarkable enhancement of NFATc1 expression, and formation of MNCs and actin ring in cells treated with RANKL and AlF4- simultaneously was identified. These results strongly suggest that activation of G-protein by AlF4- acts as a synergic factor that somehow interacts with RANKL-mediated other signals responsible for transmitting differentiation-related signals to downstream. Our results raise an important question of which Gα subunit is activated and causes the enhancement of osteoclastogenesis. There are the major four families, Gαq/11, Gαs, Gαi, and Gα12/13 whose main effector molecules are thought to be PLC, adenyly cyclase, and small GTPase families [30]. Following studies support the involvement of Gα subunits including Gαq/11, Gαs, and Gαi in osteoclastogenesis. OGR1, which is one of GPCR coupled to Gαq/11 and triggers [Ca2+]i mobilization, is reported to be involved in RANKL-induced osteoclastogenesis by modulating NFATc1 signaling pathway [11,12,31]. Continuous production of cAMP mediated by dopamine D1-like receptor, which is coupled to Gαs subunit, contributes to RANKL-induced osteoclastogenesis, whereas activation of dopamine D2-like receptor, which is coupled to Gαi subunit and inhibits adenylyl cyclase, suppresses RANKL-induced osteoclastogenesis [32]. Furthermore, critical role of CREB (cAMP response element binding protein) in RANKL-mediated osteoclastogenesis strongly supports that finely tuned cAMP production by Gαs and Gαi and Gαq/PLC/IP3/Ca2+ signaling pathway are a key modulator in osteoclastogenesis [25,33,34]. Taken together, it is convinced that modulating the activities of Gαq/11, Gαs and Gαi affects on RANKL-induced osteoclastogenesis. Since our current results clearly show that treatment of AlF4- induces [Ca2+]i mobilization in a form of oscillations and enhances RANKL-mediated osteoclastogenesis, Gαq/11 seems to dominantly be involved in RANKL-mediated osteoclastogenesis. However, due to the property of AlF4- as a universal activator of Gα subunit, it should be another important study to determine which subtype of Gα subunit governs signaling pathway related to differentiation.
Before the finding of co-stimulatory signal pathway including Ca2+ increase via DAP12 and FcRγ, it is generally accepted that TRAF6/MAPK/AP-1/NFATc1 pathway mainly dominates gene induction responsible for late-stage of osteoclastogenesis [2,35,36]. As shown in Fig. 4, RANKL is known to activate MAPK pathway, such as ERK and JNK, at early moment. Consistent with previous results, activation of ERK and JNK is appeared to be activated in response to not only co-stimulation with RANKL and AlF4- but also AlF4- alone. Although this result bring puzzling question that is subject to different interpretations with respect to the cause-effect relationship between [Ca2+]i oscillations and MAPK, we could assure that AlF4- is sufficient to activate MAPK necessary for osteoclastogenesis. One of important finding in this result is that function of G-protein responsible for differentiation into osteoclast is associated with either pathway of Ca2+ mobilization and MAPK by which enhance cell fusion and attachment of cell on bone surface. The role of GPCR in osteoclastogenesis is still uncertain though there are several reports showing involvement of signal molecules following GPCR activation in osteoclastogenesis. The other is the application of AlF4- for inducing various cell functions by activating G-protein. AlF4- acts as a messenger of false information [13] and have been often used in many laboratories to investigate their effects on various cells and tissues. Further studies are needed to clarify the potential risks for human health of long-term exposure to AlF4-. Notably, our study indicates the possibility that Gα subunits can be regarded as a target molecule necessary on modulating bone resorption. As mentioned previously, OGR1 (GPR68), which is coupled to Gαq/11 subunit and transmit signals to PLC, is one of few GPCRs that is demonstrated the roles in osteoclastogenesis. Despite of the absence of direct evidence, following studies presented the answer for the question. 1) Deficiency of OGR1 caused abnormalities in osteoclastogenesis [9], 2) RGS18, which accelerates intrinsic GTP hydrolysis on heterotrimeric G-protein alpha subunits, negatively regulates osteoclastogenesis by modulating OGR1/NFAT signaling pathway [11]. Unless RANKL directly activates Gα subunits, inhibition of G alpha subunits may not block the osteoclastogenesis. However, assuming Gα subunits is inhibited in vivo, it can cause the reduction of osteoclastogenesis and bone resorption.
In summary, present study demonstrates that AlF4- induces [Ca2+]i oscillations and MAPK, which lead to improve RANKL-mediated osteoclastogenesis by enhancing NFATc1 expression. Although not described in detail here, it is needed that further verification for signal molecules related with G-protein, such as GPCR and RGS protein, during osteoclastogenesis. With this finding, it should be a novel therapeutic target for bone-related disorders including osteoporosis and osteopetrosis.
Go to :
 XML Download
XML Download