

 PDF
PDF ePub
ePub Citation
Citation Print
Print


ABBREVIATIONS
HCC
hepatocellular carcinoma
HBV
hepatitis B virus
HCV
hepatitis C virus
CypA
cyclophilin A
Cyps
cyclophilins
CsA
cyclosporine A
PPIase
peptidyl prolyl isomerase
ER
endoplasmic reticulum
MPTP
mitochondrial permeability transition pore
HIV-1
human immunodeficiency virus-1
IFN
interferon
IL
interleukin
INTRODUCTION
Cyclophilins (Cyps) constitute a subgroup of large family proteins called immunophilins. Cyps are well conserved and are found throughout evolution from E. coli to humans. Cyclosporine A (CsA) is a reversible inhibitor for all Cyps and works at nanomolar concentrations [1,2]. Cyps possess a peptidyl prolyl isomerase (PPIase) that catalyzes the conversion between cis-trans isomers of proline on the peptide. PPIases are regarded as a chaperone [3] and work with other chaperone proteins such as the heat shock protein and protein disulfide isomerases [4,5]. PPIases not only function in folding of de novo synthesized proteins but also accelerate slow steps in the refolding of denatured proteins [6]. There are 18 human Cyp family proteins which include Cyp isoforms such as CypA, CypB, CypC, CypD, CypE, Cyp40, Cyp20 and CypNK. Although complete isoform characterization is required to define precise functions for the individual Cyp proteins, multiple Cyps display separate functions by localizing through unique domains for particular cellular compartments such as CypA in the cytosol and nucleus, CypB in the endoplasmic reticulum (ER), CypD in mitochondria. The substrates or specific roles of individual Cyps in their subcellular locales or during critical cellular processes are still largely unknown. Chaperone activity of Cyps has been demonstrated first time when inhibition of the PPIase significantly delays maturation of collagen [7,8]. Cyps are also involved in repairing stress-induced damaged proteins like other molecular chaperon proteins. Environmental stresses can induce some Cyp expressions to carry out the repair function [9,10]. Recently, human Cyps emerge to be recognized as an important protein by participating in important physiological tasks like muscle differentiation [11] and immune response [12].
CypA forms a ternary complex with CsA, which inhibits calcineurin phosphatase activity and exerts an immunosuppressive action. CypB interacts with Stat3 and influences its transcriptional activity [13]. The prevention of mitochondrial permeability transition pore (MPTP) opening and anti-apoptotic effect by CsA is from its CypD inhibition. CypD is found in mitochondria and plays a role in apoptosis, being a member of the MPTP complex with ANT, VDAC, and hexokinase [14,15]. Cyp20 is important for folding of mitochondrial proteins as well as for transporting newly synthesized proteins between intracellular compartments [16,17]. In addition to the intracellular functions, extracellular Cyps such as CypA and CypB may contribute to intercellular communication inducing signaling responses in target cells [18]. CypB as well as CypA can be secreted from cells and those secretory Cyps can be a chemoattractant to immune cells [19,20]. It is reported that the secreted forms of CypA and CypB may function as a ligand and activate signal transduction pathways in various cell types [21]. CD147, an Ig-like membrane receptor, is involved in the signaling and has been proposed as a receptor for both CypA and CypB [22,23]. This binding of CypA to CD147 can become a pathological factor for rheumatoid arthritis [24]. Furthermore, CypA can be secreted in response to oxidative stress, which triggers the pathogenesis of vascular diseases [25].
CypA, found in almost all tissues, is the most abundant among the Cyps. The cytosolic CypA concentration may account for as much as 0.1~0.4% of the total protein in a cell [26]. CypA as a multifunctional chaperone has been demonstrated to function in intracellular trafficking [27], signal transduction and transcription regulation [28]. CypA's function in pathophysiological conditions has currently attracted attentions of researchers in the field of oncogenic development, cardiovascular diseases as well as viral infections [29,30]. Overexpression of CypA often correlates with severity of cancers including pancreatic and hepatic cancers, suggesting its role in the tumor malignancy. CypA can help infections of viruses including the clinically important ones such as human immunodeficiency virus-1 (HIV-1) [31], HBV and HCV [32,33].
The role of CypA in helping viral infection has primarily been studied in the case of HIV infection. CypA helps HIV-1 assembly by binding to HIV-1 capsid proteins [34]. Cyps, especially CypA, has been studied extensively and found to play essential roles during HCV infection. Recent studies have demonstrated that CypA is the one to interact with HCV nonstructural protein NS5A and essential for the viral genome replication although interaction of CypB and RNA polymerase, NS5B, was proposed first [35]. Likewise, the three nonimmunosuppressive Cyp inhibitors (NIM811, SCY-635, and Alisporivir) that are now under clinical trials on HCV infections are believed to be effective through CypA inhibition [36]. It is found that infections with hepatotropic viruses are linked to the majority of cases of HCC and in fact, chronic infections with HBV and HCV account for up to 80% of HCCs [37,38]. HCC development follows the sequential steps of chronic inflammation, liver regeneration, fibrosis, and cirrhosis [39,40]. While the molecular mechanisms of hepatocyte transformation are not fully understood, it is believed to involve alterations of multiple cellular signal transduction pathways and changes in micro (mi)RNAs and cytokines. Inflammatory cytokines appear to be an important component of the immune system and play an important role in promoting HCC development. In this review, the role of CypA in HCV infection and subsequent HCC development will be discussed.
ROLE OF HCV IN THE DEVELOPMENT OF HCC
HCC is one of the most common cancers and the third leading cause of cancer-associated death [41]. It takes 30~40 years for HCV-infected patients to develop HCC. Recent investigations of HCV reveal several cellular signal transduction pathways that are altered by HCV infections [39]. HCV possesses four structural proteins (core, E1, E2, and p7) and six nonstructural (NS2, NS3, NS4A, NS4B, NS5A, and NS5B) viral proteins [42]. The HCV non-structural proteins play important roles in virus-host interactions and thus may help avoid the immune response. HCV translation occurs on the rough ER and host signal peptidases play important roles in processing the structural proteins. Core composes the viral nucleocapsid by interactions with the HCV RNA. The E1 and E2 are envelope glycoproteins which form heterodimers in the ER. E1 and E2 together may mediate entry of the viral particles into cells. Viroporin is the name for p7, a small transmembrane protein with ion channel activity. However, the exact role of p7 in the HCV life cycle is poorly understood. NS2 is a cysteine protease while NS3 is a complex serine protease with helicase and adenosine triphosphatase activities. Importantly, the nonstructural proteins are also required for RNA replication and these NS2 and NS3 viral proteases cleave HCV polyprotein, generating NS4A, NS4B, NS5A, and NS5B. NS3 with NS4A, an NS3 cofactor, in necessity form an NS3/NS4A complex and embedded in the ER and the mitochondrial outer membrane. The NS3A/4A complex is crucial in viral resistance to the innate immune response by inactivation of the interferon (IFN) regulatory factor 3 and cleavage of the Toll-like receptor 3 adaptor protein, TRIF. NS4B is also required for HCV replication and NS5A is involved in HCV replication and virion assembly. NS5B is an RNA-dependent RNA polymerase. HCV core, NS3 and NS5A proteins are known to interrupt p53 actions and some HCV structural proteins may have a direct oncogenic role [43].
In addition to the viral proteins, modulation in the host factors such as miRNA and cytokines also play important roles in viral pathogenesis. Connection of miRNA functions to HCV-related hepatocarcinogenesis draws an attention because modulation of miRNA profiles can be associated with hepatocarcinogenesis [44]. miRNAs result in gene silencing via translational repression or mRNA degradation [45,46]. Many miRNAs that are differentially expressed in HCC appear to play roles in both viral replication and HCC development [47]. miRNA-122 is known to enhance HCV RNA translation [48-51]. miRNA-122 binds to the 5' UTR of HCV and is required for HCV replication in hepatocytes [52]. Besides, downregulation of miRNA-145 and miRNA-199b as well as upregulation of miRNA-224 are detected during HCC development [53]. Downregulation of miRNA-26 is also detected in patients with HCC [54]. Since expression profiles of miRNAs are modulated by HCV and are found to be altered in human HCC, miRNAs may become a diagnostic and prognostic tool and potential therapeutic targets for the treatment of HCV-associated HCC.
HCV completes its life cycle in the cytoplasm and does not integrates its RNA genome in the host DNA. Therefore, HCV-related HCC chromosomal aberrations cannot be found and genomic instability and oncogene activation are not driven by insertional mutagenesis [55]. Instead, HCV infection appears to promote carcinogenesis through the induction of chronic inflammatory states [56]. Cirrhosis is developed among the 25% of chronically HCV-infected patients and ultimately may lead the disease state into HCC [57]. Therefore, development of HCV-associated HCC is closely associated with liver inflammation, fibrosis and chirrohsis after chronic infection. During chronic HCV infection, HCC accompanies altered intracellular signaling pathways. HCV infection influences the host immune system, modulating cytokines in the microenvironment, further promoting HCV infection, and possibly leading to HCC development [58]. Chronic infection can signify persistent cytokine production and influence the cytokine balance. Cytokines play important roles against viral infections and viral polypeptides and interact with cells involved in innate and adaptive immunity [59]. While many cytokines have pleiotropic effects, they can be categorized as proinflammatory or anti-inflammatory. CD4 Th cells secrete mainly interleukin (IL) cytokines. IL cytokines can be divided into Th1 cytokines like IL-1α, IL-1β, IL-2, IL-12p35, IL-12p40, IL-15, TNF-α and IFN-γ that generally induces proinflammatory responses and the other Th2 cytokines like IL-4, IL-8, IL-10, and IL-5 that induce anti-inflammatory responses. Aberrant immune responses after chronic infection accompanying imbalance of proinflammatory Th1 and anti-inflammatory Th2 cytokines can initiate carcinogenic cascade in liver. The proinflammatory Th1 and anti-inflammatory Th2 cytokine balances are impaired in the microenvironment of HCC patients. The pro-and anti-inflammatory cytokines that are deviated from normal immune responses contribute to chronic infection. TNF-α is the most important mediator of liver inflammation, which has multiple functions in the development of HCC [60]. TNF-α together with tumor growth factor-β, IL 1, and IL-6 is an important factor, inducing the production of fibrogenic factors and therefore contributing to fibrosis in chronic liver disease [61]. The proinflammatory cytokine IFN-α is currently used for treatment of chronic viral hepatitis and to prevent development of HCC while proinflammatory TNF-α is associated with an increase in cell-cycle progression [62]. Cytotoxic effect of TNF-α is inhibitory to the HCC development but TNF-α can help hepatocytes to be carcinogenic. The repetitive cell death and regeneration of hepatocytes cause accumulation of mutations [63-65]. IL-8, anti-inflammatory cytokine, shows an angiogenic function and can accelerate the proliferation of various HCC cell lines [66]. Increased IL-10 expression is associated with the development of HCC [67-69]. HCV contributes to destroy cellular immunity by inducing IL-10. Interestingly, obesity-induced HCC undergoes similar processes which include lipid accumulation and chronic inflammation with modulations in cytokines and adipokines [70]. Among the cytokines, IL-6 is a marker for obesity-associated inflammation. The activation of genes involved in the IL-6 signaling pathway has been reported during the development of HCC [71]. Activation of TNF-α has an important factor for the inflammatory process and TNF-α levels correlate with the degree of inflammation [72,73]. HCV core proteins can directly modulate several cytokines. NS5A upregulates the expression of TGF-β as well as TGF-β2 and VEGF proteins [74,75], and modulates TGF-β signaling through interaction with TGF-β receptor I [76,77]. Virus proteins such as HCV p21core, E2 envelope protein and nonstructural NS5A may directly alter cytokine productions in the liver. HCV p21core decreases the production of IFN-γ and IL-2 by suppressing HCV-specific CTL responses [78]. Furthermore, p21core directly interacts with TNF-type receptors and p21core increases Fas-mediated apoptosis and liver infiltration of peripheral T cells [79,80]. The E2 protein inhibits NK cells and NS5A inhibit IFN-α-induced protein kinase [81,82]. IL-10 is highly expressed in HCC tumors, suggesting that high levels of IL-10 or other Th2 cytokines may be associated with HCC progression [83-87]. Proinflammatory Th1 cytokines such as IL-1β, IL-15, IL-18, TNF-α, IL-12 and IL-2 expression are also known to be elevated in HCC patients [88-90]. Increased TNF-α/IL-10 ratio is associated with cirrhosis and HCC and thus high TNF-α/IL-10 ratio may show an increased risk to develop severe hepatic disease [91]. All these suggest the importance of the microenvironmental factors such as host-viral interactions in triggering HCC after chronic inflammation. In addition, HBV- and HCV-encoded proteins can directly act to alter the hepatocyte physiology and hepatocellular signal transduction pathways [92,93]. Therefore, hepatocarcinogenesis may include both indirect, namely inflammation-mediated, and direct HBV or HCV protein-induced alterations in hepatocyte physiology.
CYPA FUNCTION IN CANCER
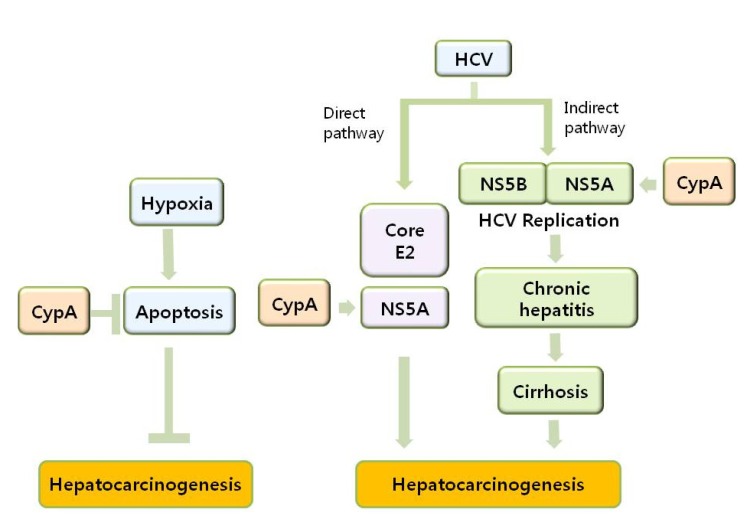
The inflammatory responses after viral infections such as destruction and regeneration of hepatocytes and expression of HCV-encoded proteins will contribute to hepatocyte transformation. The mechanisms of CypA as a host factor during HCV infection and HCC development include anti-apoptotic effect as well as direct and indirect pathways (Fig. 1). CypA, predominantly expressed in human cells, was initially found to be upregulated in hepatocellular carcinoma [94,95]. Overexpression of various Cyps has been repeatedly observed in many cancers and their expression levels and patterns in different cancer types have been considerably well documented. However, the process of how each Cyps participate in the biochemical mechanisms of tumourigenesis is far from being understood. CypA has been reported to be overexpressed in diverse cancer types of small cell lung cancer [96,97], pancreatic cancer [98-101], breast cancer [102,103], colorectal cancer [104-106], squamous cell carcinoma [107,108], melanoma [109], and glioblastoma multiform [110]. Upregulation of CypA in small cell lung cancer stimulates cancer cell growth [111,112]. Subsequently, Qi et al. suggested that CypA may positively function during malignant transformation of esophageal squamous cells [108]. The fold increase of CypA can be found up to 5 times and CypA is also involved in metastasis [109]. Since the proteins with chaperone activity are often stress proteins that protect cells from stress-induced protein damage or denaturation, CypA up-regulation in response to stresses may be regarded naturally as other chaperones. Studying if the CypA up-regulation in cancer is controlled by the same regulatory mechanisms of stress induction can help to understand CypA-involved cancer development. Upregulation of CypA in cancers by hypoxic inducible transcription factor-1α (HIF-1α) has been reported. HIF-1α induces CypA expression by HIF-1α binding to hypoxia response elements in the CypA promoter region under hypoxic conditions [113]. CypA can also be overexpressed during p53-induced apoptosis [114,115]. It will be intriguing to know if interaction of CypA with p53 may lead to stabilization of p53.
Clinical implications of the overexpressed CypA in cancers are currently investigated with an expectation of CypA being a therapeutic target. During development of solid tumors, reactive oxygen species are continuously generated in the central hypoxic region. CypA may exert antioxidant effects on the hypoxic regions in solid tumors through its PPIase activity [11]. Overexpression of CypA in cancer cells renders resistance to hypoxia- and cisplatin-induced cell death in a p53 independent manner [113]. CypA are also regulated by chemotherapeutic agents; for example, cisplatin causes CypA overexpression and induces resistance to several chemotherapeutic agents [98]. Treatment with chemotherapeutic agents such as celecoxib and 5-fluorouracil, on the contrary, is reported to lower CypA expression [105,106].
Human Cyps appear to show pleiotropic effects via conformational interconversions of prolyl cis-trans isomerization on the backbones of key proteins in signaling pathways. Recent studies indicate that PPIases utilize isomerization catalysis on the substrate proteins under both physiological and pathophysiological conditions. Then, the development of isoenzyme-, tissue- or organelle-specific PPIase inhibitors will open a new road for cancer intervention strategies.
CYPA AS A THERAPEUTIC TARGET FOR HCC
Targeting CypA for cancer therapeutics has recently begun to emerge because CypA overexpression is often found in many different cancer types and CypA appears to be involved in malignant transformation in some cancer types. In parallel, CypA inhibitors show antiviral activity and anti HCC effect as replication of HCV RNA genome depends on the expression of CypA. Pegylated IFN-α and ribavirin that directly target the virally-encoded proteins are a current treatment regimen for chronic HCV infection [116]. Viral protein inhibitors, however, show a limited response with severe side effects and also develop high viral resistance [117-119]. The CypA inhibitors are advantageous because host targeting can provide a higher barrier to resistance than viral inhibitors. Both CypA and CypB are important host factors for the HCV replication process since interactions of both CypA with NS5A and CypB with the HCV RNA-dependent RNA polymerase NS5B contribute to form a multi-protein complex [120]. Treatment of CypA inhibitors cause dissociation from the protein complex that is formed between CypA and NS5A [121-123]. The dissociation by CypA inhibitors decreases the affinity of NS5A for viral RNA and inhibits viral replication [124-128]. CsA was first tested to be effective against HCV and afterwards, Cyp inhibitors without immunosuppressive activity were developed for treatment of HCV infection [129-133]. CsA derivatives exert anti-HCV activity by inhibiting host CypA and CypA has not been implicated in the mechanism of action of either pegylated IFN-α or ribavirin [134]. Therefore, synergistic effects against HCV of are expected when CypA inhibitors and conventional drugs are used in combination. Three CsA derivatives, NIM811, SCY-635, and Alisporivir have shown promising anti-HCV responses in pre-clinical and clinical trials and the resistance level was substantially lower than viral protein inhibitors [135].
The anti-HCV mechanisms of action of these CypA inhibitors are not clearly defined yet. However, recent studies unravel some fundamental findings. The inhibitors inhibit PPIase activity of CypA and the isomerase active site of CypA is critical for hepatitis C virus replication [136-138]. CypA binds to prolines located in the domain II of the HCV NS5A protein and Cyp inhibitors interfere the Cyp A-NS5A interactions in a dose-dependent manner [139-141]. While PPIase activity of CypA suggests the function of CypA in structural interconversion of the interacting protein, how the interaction between CypA and NS5A influences HCV replication remains to be elucidated. With the finding that CypA specifically promotes NS5A binding to viral RNA, it has been suggested that cis-trans isomerization of NS5A proline bonds by CypA enhances NS5A binding to viral RNA and somehow facilitates HCV replication [142-144]. Since NS5A binds the HCV polymerase NS5B directly, interaction of CypA with NS5A may affect NS5B polymerase activity. In addition, interaction of CypB with NS5B is important for HCV replication and the anti-HCV effects of Cyp inhibitors may be the outcome of blocking modulations of both NS5A and NS5B by CypA and CypB [145,146].
NIM811 was discovered as 2-5-fold higher binding affinity to Cyps than CsA and inhibited HCV replication in vitro with an EC50 of 0.12 µM in the absence of human serum and 0.63 µM in the presence of 40% human serum. NIM811 was well tolerated and there appeared to be no drug-related adverse event during evaluation. NIM811 was limited by its significant side effect in decreasing platelet [147]. Alisporivir is another CsA analogue without immunosuppressive action [147]. Alisporivir is more potent than NIM811 in inhibiting HCV with an EC50 of 0.045 µM and a serum (100%) adjusted EC50 of 0.33 µM [148]. While it was initially developed for HIV-1, Alisporivir displayed high antiviral activity against HCV RNA [149]. In behalf of its higher barrier to resistance, an IFN-free regimen was carried out to treat IFN non-responders in a Phase II trial and HCV became undetectable in 75.4% of the patients [150]. Alisporivir is the first time trial for a host-targeting antiviral alone and effective both in vitro and in patients, being considered as an ideal candidate inhibitor as an IFN-free, all-oral regimen [151]. The main side effect was a jaundice, however, without any increase in ALT, AST or γ-GT, or with hemolysis [152]. Unfortunately, six cases of severe pancreatitis during phase III trial were reported [153]. SCY-635 was another CsA analog with an EC50 of 0.03 µM [154]. It showed a different side-effect of elevated serum creatinine phosphokinase and ALT/AST, but there was no signs of bilirubin increase or platelet decrease. Recent clinical studies suggest antiviral combination therapies toward IFN-free and ribavirin-free regimens and Alisporivir possessing prominent features as described is under clinical trials for combination with specific direct-acting antiviral agents.
SUMMARY
HCV infections frequently advance into the development of HCC and considerable evidence suggests that 1) inflammatory responses, 2) associated destruction and regeneration of hepatocytes, and 3) activities of HCV-encoded proteins contribute to hepatocyte transformation. Viral proteins and inflammatory responses are clearly associated with the alterations in hepatocyte physiology and the liver environment. However understanding the mechanisms involved is still incomplete and further studies warrant effective strategies for treating HCV infections and preventing HCC. Current knowledge of the proteomics of HCV-induced liver disease is still developing and protein profiles from recent discoveries will be important for diagnosis and treatment of HCV-induced HCC. In the meantime, targeting CypA for cancer therapeutics has been earnestly sought because CypA overexpression is common in malignant cancer cells. Simultaneously, CypA inhibitors can show antiviral activity and thus be benefited as HCC therapeutic agents. CsA suppresses HCV genome replication in HCV replicon cells and human hepatocytes infected with HCV. However, the inhibition of the phosphatase calcineurin by CsA and CypA complex display immunosuppresive action which hinders its use as an antiviral agent. Thereupon, specific CsA derivatives devoid of immune suppressive effects such as NIM811, SCY-635, and Alisporivir have been developed for HCV treatment. In fact, SCY-635 and Alisporivir have emerged as powerful antiviral agents since they inhibit the in vitro replication of HCV subgenomic replicons. Furthermore, clinical use of Alisporivir as an anti-HCV agent has been confirmed in patient studies. In conclusion, the CypA inhibitors may represent a candidate as therapeutic agents for the HCV-induced HCC by preventing both HCV infection and hepatocarcinogenesis.
 XML Download
XML Download